

Sulfenome mining in Arabidopsis thaliana
- PMID: 25049418
- PMCID: PMC4128149
- DOI: 10.1073/pnas.1411607111
Sulfenome mining in Arabidopsis thaliana
Abstract
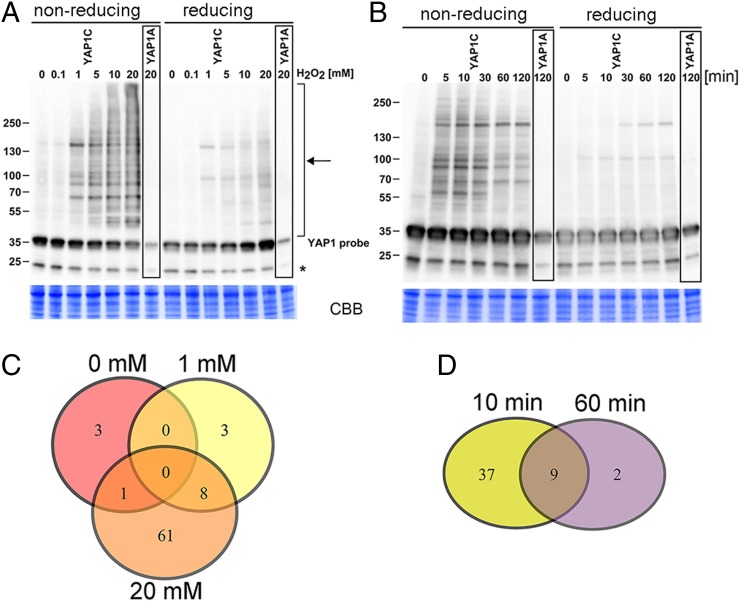
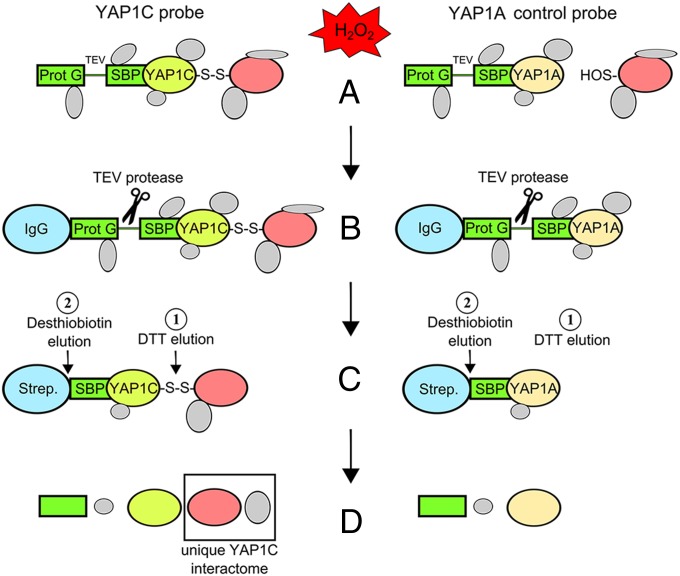
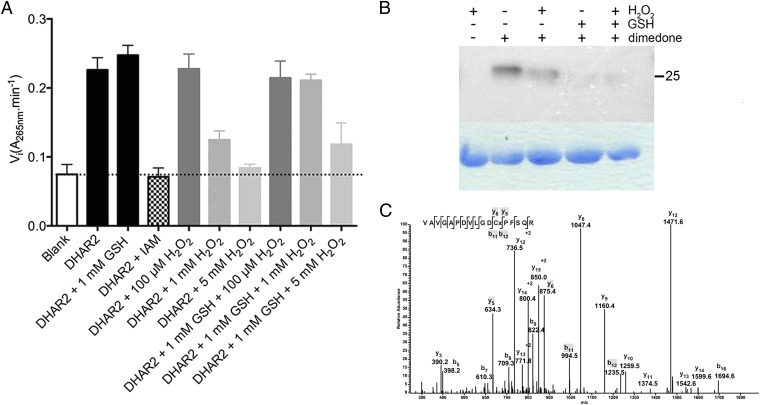
Reactive oxygen species (ROS) have been shown to be potent signaling molecules. Today, oxidation of cysteine residues is a well-recognized posttranslational protein modification, but the signaling processes steered by such oxidations are poorly understood. To gain insight into the cysteine thiol-dependent ROS signaling in Arabidopsis thaliana, we identified the hydrogen peroxide (H2O2)-dependent sulfenome: that is, proteins with at least one cysteine thiol oxidized to a sulfenic acid. By means of a genetic construct consisting of a fusion between the C-terminal domain of the yeast (Saccharomyces cerevisiae) AP-1-like (YAP1) transcription factor and a tandem affinity purification tag, we detected ∼ 100 sulfenylated proteins in Arabidopsis cell suspensions exposed to H2O2 stress. The in vivo YAP1-based trapping of sulfenylated proteins was validated by a targeted in vitro analysis of dehydroascorbate reductase2 (DHAR2). In DHAR2, the active site nucleophilic cysteine is regulated through a sulfenic acid-dependent switch, leading to S-glutathionylation, a protein modification that protects the protein against oxidative damage.
Keywords: cysteine oxidation; oxidative stress; redox regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Garavelli JS. The RESID Database of Protein Modifications as a resource and annotation tool. Proteomics. 2004;4(6):1527–1533. - PubMed
-
- Roos G, Messens J. Protein sulfenic acid formation: From cellular damage to redox regulation. Free Radic Biol Med. 2011;51(2):314–326. - PubMed
-
- Ma L-H, Takanishi CL, Wood MJ. Molecular mechanism of oxidative stress perception by the Orp1 protein. J Biol Chem. 2007;282(43):31429–31436. - PubMed
-
- Mou Z, Fan W, Dong X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell. 2003;113(7):935–944. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
