

Uric acid accumulation in an Arabidopsis urate oxidase mutant impairs seedling establishment by blocking peroxisome maintenance
- PMID: 25052714
- PMCID: PMC4145134
- DOI: 10.1105/tpc.114.124008
Uric acid accumulation in an Arabidopsis urate oxidase mutant impairs seedling establishment by blocking peroxisome maintenance
Abstract
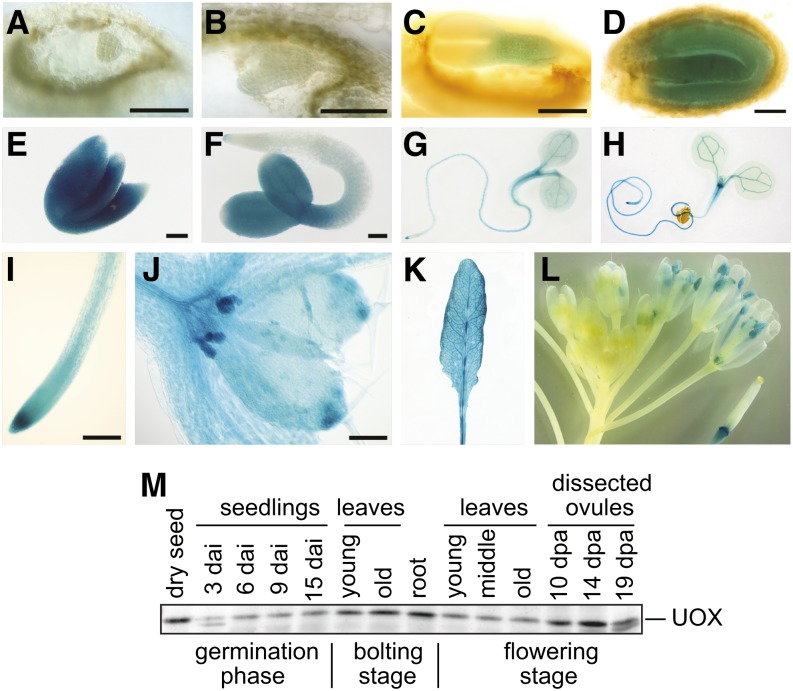
Purine nucleotides can be fully catabolized by plants to recycle nutrients. We have isolated a urate oxidase (uox) mutant of Arabidopsis thaliana that accumulates uric acid in all tissues, especially in the developing embryo. The mutant displays a reduced germination rate and is unable to establish autotrophic growth due to severe inhibition of cotyledon development and nutrient mobilization from the lipid reserves in the cotyledons. The uox mutant phenotype is suppressed in a xanthine dehydrogenase (xdh) uox double mutant, demonstrating that the underlying cause is not the defective purine base catabolism, or the lack of UOX per se, but the elevated uric acid concentration in the embryo. Remarkably, xanthine accumulates to similar levels in the xdh mutant without toxicity. This is paralleled in humans, where hyperuricemia is associated with many diseases whereas xanthinuria is asymptomatic. Searching for the molecular cause of uric acid toxicity, we discovered a local defect of peroxisomes (glyoxysomes) mostly confined to the cotyledons of the mature embryos, which resulted in the accumulation of free fatty acids in dry seeds. The peroxisomal defect explains the developmental phenotypes of the uox mutant, drawing a novel link between uric acid and peroxisome function, which may be relevant beyond plants.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures







References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
-
- Angelovici R., Galili G., Fernie A.R., Fait A. (2010). Seed desiccation: A bridge between maturation and germination. Trends Plant Sci. 15: 211–218. - PubMed
-
- Brychkova G., Alikulov Z., Fluhr R., Sagi M. (2008). A critical role for ureides in dark and senescence-induced purine remobilization is unmasked in the Atxdh1 Arabidopsis mutant. Plant J. 54: 496–509. - PubMed
-
- Chia T.Y.P., Pike M.J., Rawsthorne S. (2005). Storage oil breakdown during embryo development of Brassica napus (L.). J. Exp. Bot. 56: 1285–1296. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
