

Long-term evolution of nucleotide-binding site-leucine-rich repeat genes: understanding gained from and beyond the legume family
- PMID: 25052854
- PMCID: PMC4149708
- DOI: 10.1104/pp.114.243626
Long-term evolution of nucleotide-binding site-leucine-rich repeat genes: understanding gained from and beyond the legume family
Abstract
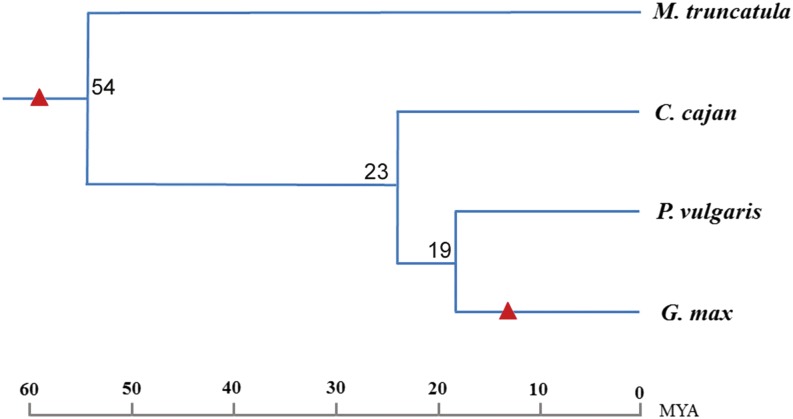
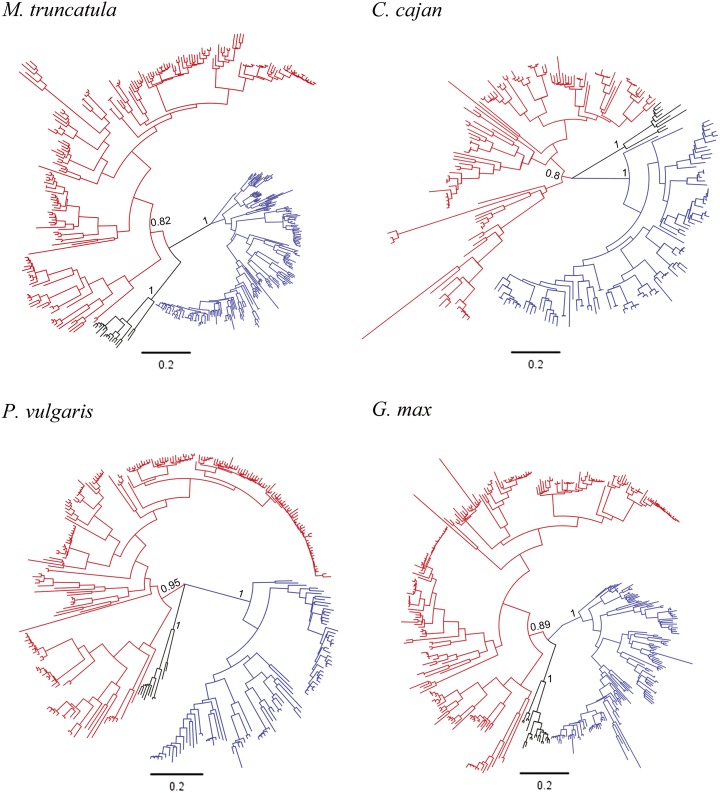
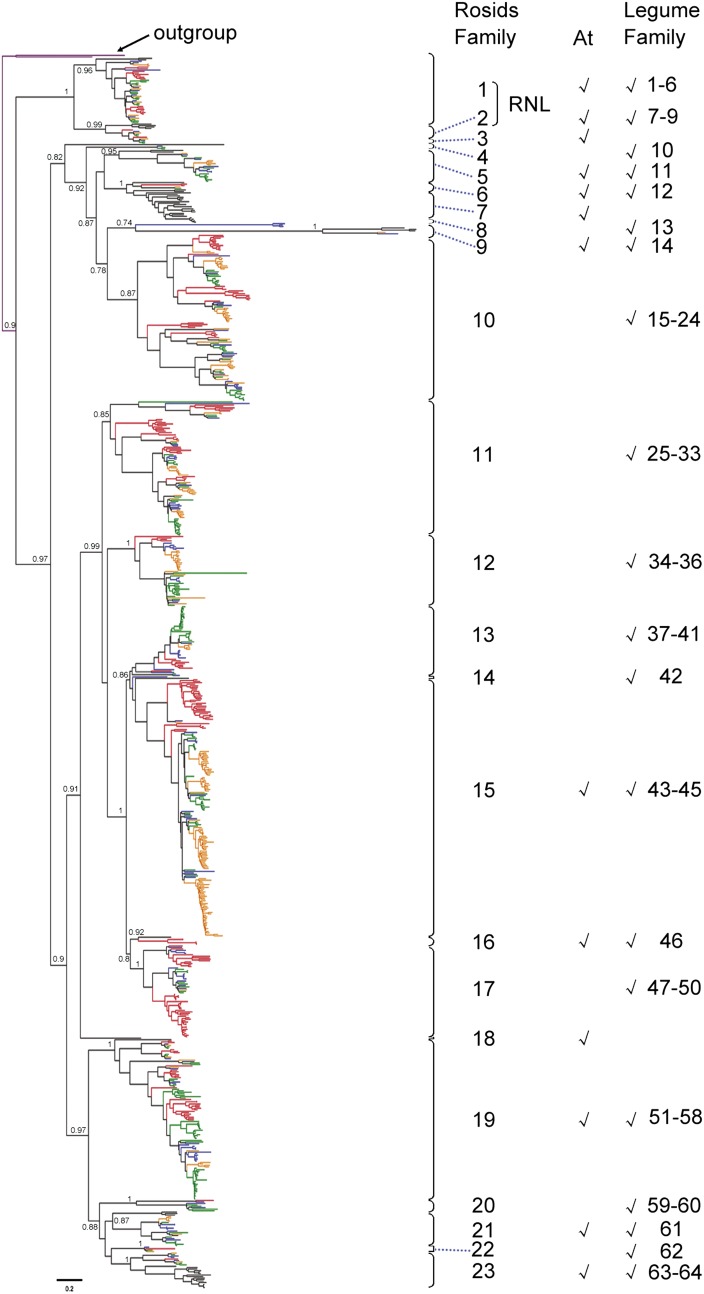
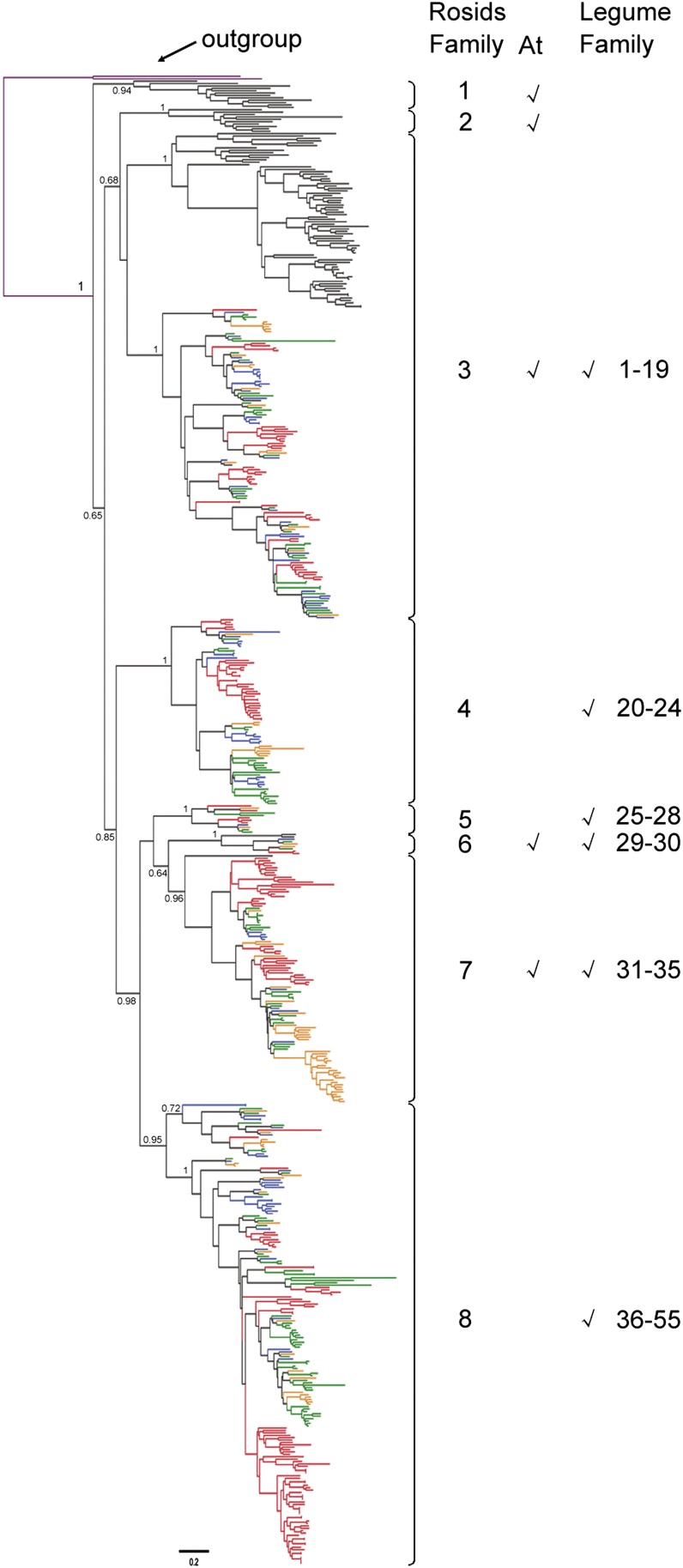
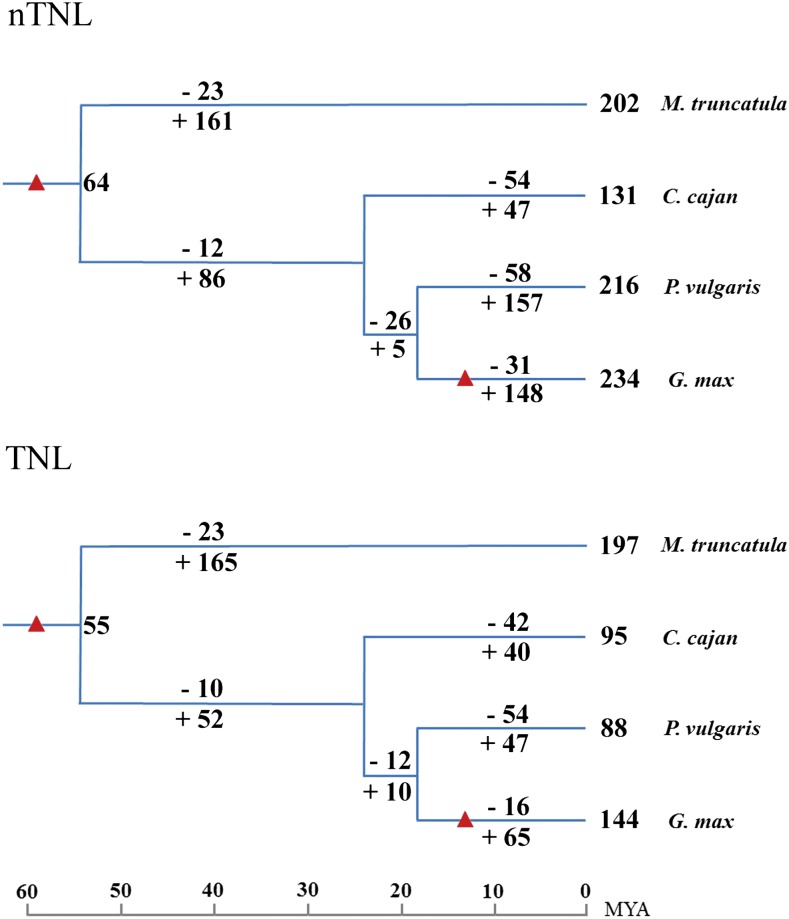
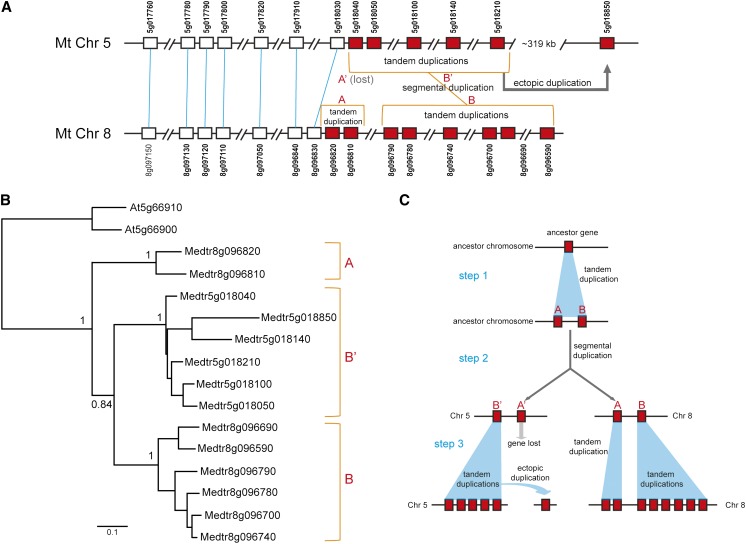
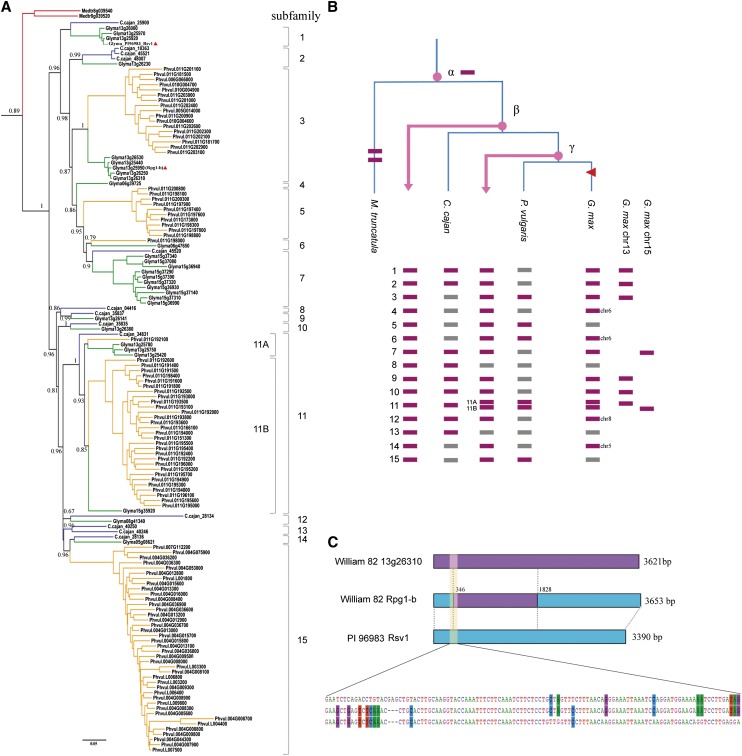
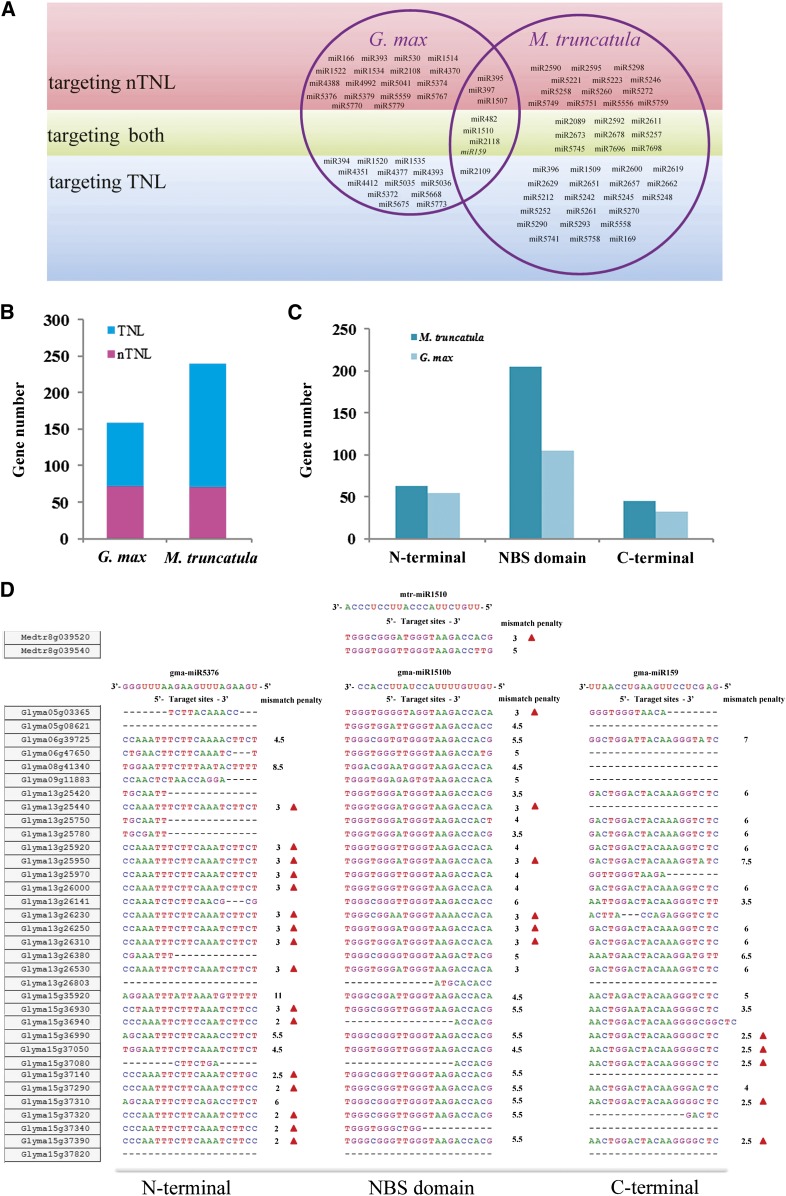
Proper utilization of plant disease resistance genes requires a good understanding of their short- and long-term evolution. Here we present a comprehensive study of the long-term evolutionary history of nucleotide-binding site (NBS)-leucine-rich repeat (LRR) genes within and beyond the legume family. The small group of NBS-LRR genes with an amino-terminal RESISTANCE TO POWDERY MILDEW8 (RPW8)-like domain (referred to as RNL) was first revealed as a basal clade sister to both coiled-coil-NBS-LRR (CNL) and Toll/Interleukin1 receptor-NBS-LRR (TNL) clades. Using Arabidopsis (Arabidopsis thaliana) as an outgroup, this study explicitly recovered 31 ancestral NBS lineages (two RNL, 21 CNL, and eight TNL) that had existed in the rosid common ancestor and 119 ancestral lineages (nine RNL, 55 CNL, and 55 TNL) that had diverged in the legume common ancestor. It was shown that, during their evolution in the past 54 million years, approximately 94% (112 of 119) of the ancestral legume NBS lineages experienced deletions or significant expansions, while seven original lineages were maintained in a conservative manner. The NBS gene duplication pattern was further examined. The local tandem duplications dominated NBS gene gains in the total number of genes (more than 75%), which was not surprising. However, it was interesting from our study that ectopic duplications had created many novel NBS gene loci in individual legume genomes, which occurred at a significant frequency of 8% to 20% in different legume lineages. Finally, by surveying the legume microRNAs that can potentially regulate NBS genes, we found that the microRNA-NBS gene interaction also exhibited a gain-and-loss pattern during the legume evolution.
© 2014 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Andolfo G, Sanseverino W, Rombauts S, Van de Peer Y, Bradeen JM, Carputo D, Frusciante L, Ercolano MR. (2013) Overview of tomato (Solanum lycopersicum) candidate pathogen recognition genes reveals important Solanum R locus dynamics. New Phytol 197: 223–237 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
