

Neuronal glycogen synthesis contributes to physiological aging
- PMID: 25059425
- PMCID: PMC4331761
- DOI: 10.1111/acel.12254
Neuronal glycogen synthesis contributes to physiological aging
Abstract
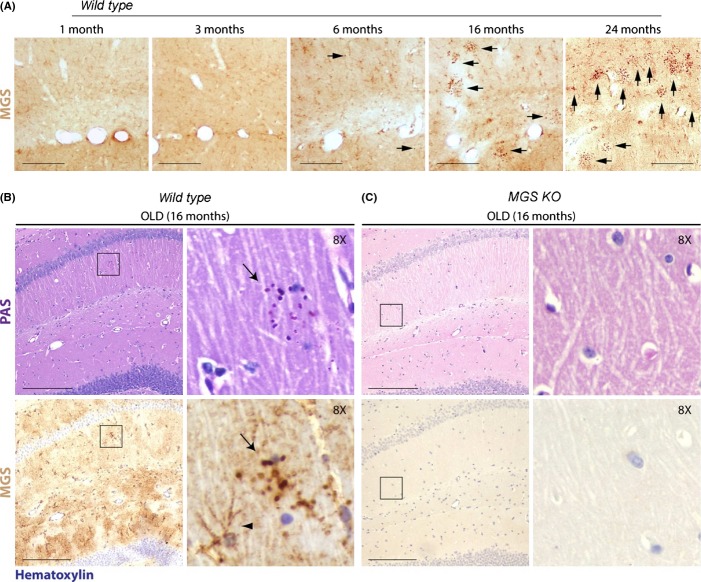
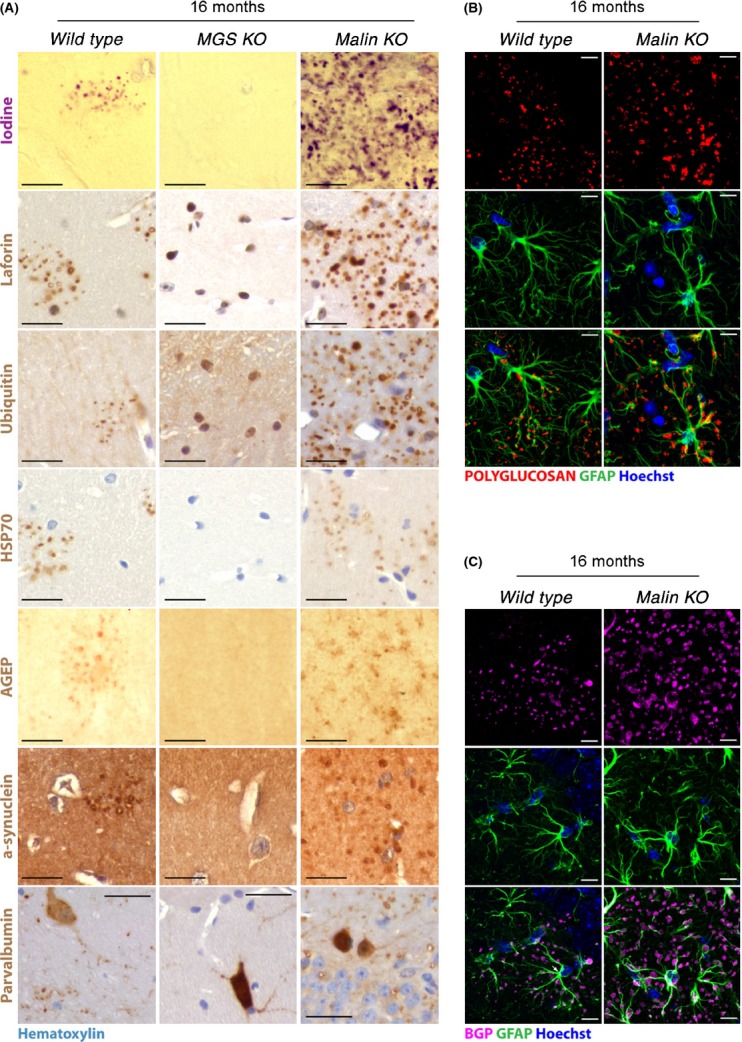
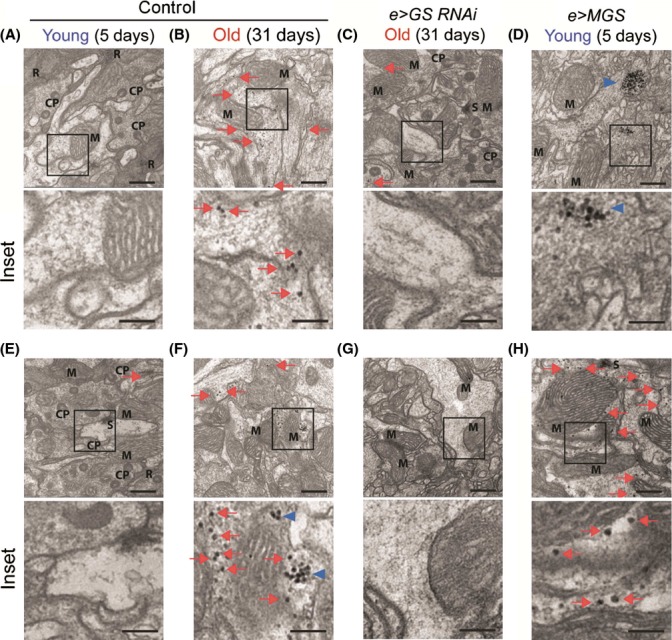
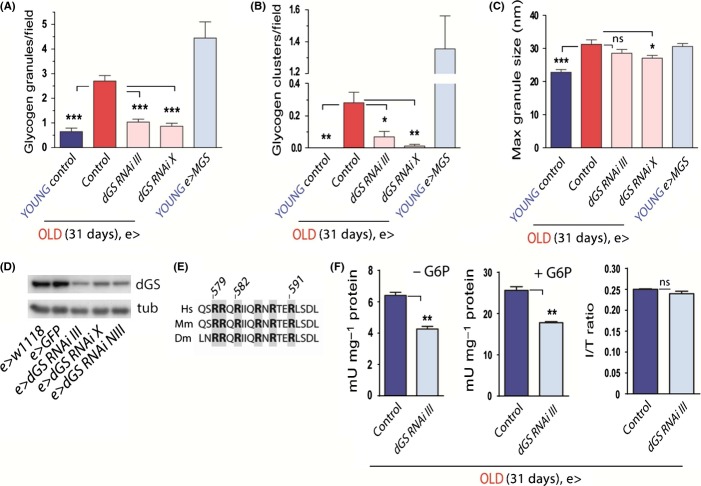
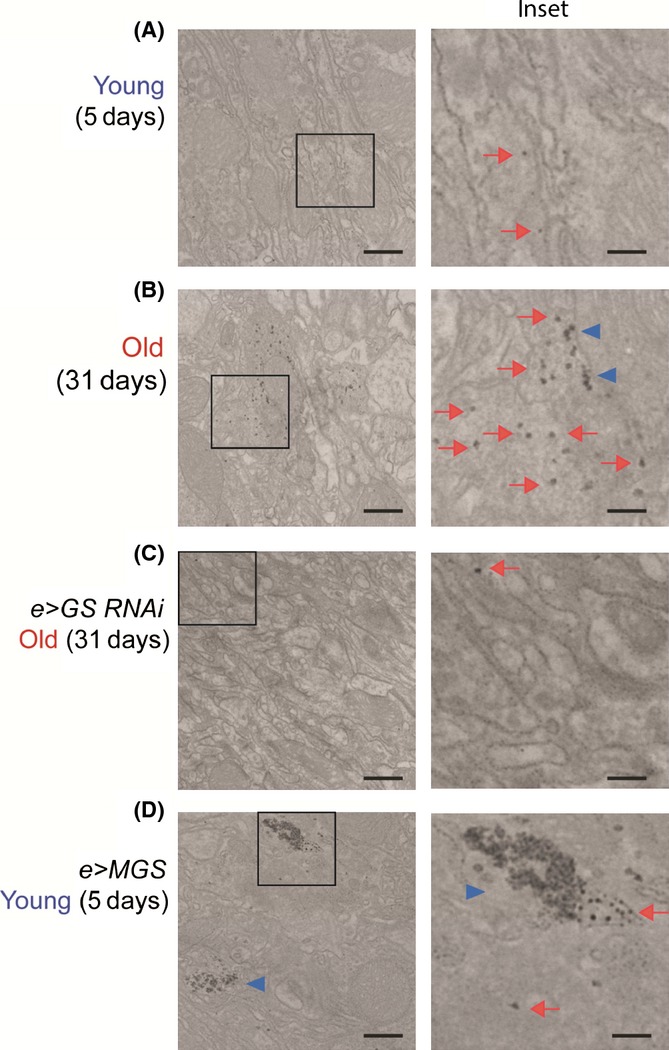
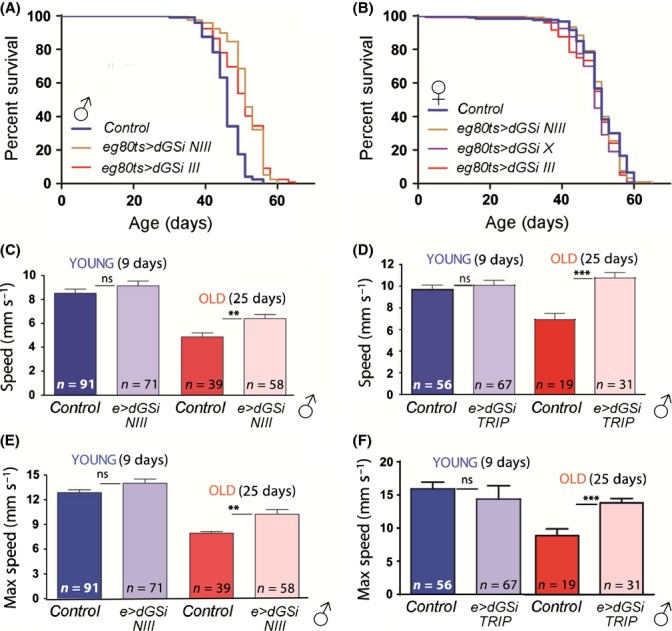
Glycogen is a branched polymer of glucose and the carbohydrate energy store for animal cells. In the brain, it is essentially found in glial cells, although it is also present in minute amounts in neurons. In humans, loss-of-function mutations in laforin and malin, proteins involved in suppressing glycogen synthesis, induce the presence of high numbers of insoluble polyglucosan bodies in neuronal cells. Known as Lafora bodies (LBs), these deposits result in the aggressive neurodegeneration seen in Lafora's disease. Polysaccharide-based aggregates, called corpora amylacea (CA), are also present in the neurons of aged human brains. Despite the similarity of CA to LBs, the mechanisms and functional consequences of CA formation are yet unknown. Here, we show that wild-type laboratory mice also accumulate glycogen-based aggregates in the brain as they age. These structures are immunopositive for an array of metabolic and stress-response proteins, some of which were previously shown to aggregate in correlation with age in the human brain and are also present in LBs. Remarkably, these structures and their associated protein aggregates are not present in the aged mouse brain upon genetic ablation of glycogen synthase. Similar genetic intervention in Drosophila prevents the accumulation of glycogen clusters in the neuronal processes of aged flies. Most interestingly, targeted reduction of Drosophila glycogen synthase in neurons improves neurological function with age and extends lifespan. These results demonstrate that neuronal glycogen accumulation contributes to physiological aging and may therefore constitute a key factor regulating age-related neurological decline in humans.
Keywords: Drosophila; aging; corpora amylacea; glycogen; protein aggregation; stress response.
© 2014 The Authors. Aging cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures
Similar articles
-
Glycogen in Astrocytes and Neurons: Physiological and Pathological Aspects.Adv Neurobiol. 2019;23:311-329. doi: 10.1007/978-3-030-27480-1_10. Adv Neurobiol. 2019. PMID: 31667813 Free PMC article. Review.
-
Brain glycogen in health and disease.Mol Aspects Med. 2015 Dec;46:70-7. doi: 10.1016/j.mam.2015.08.007. Epub 2015 Sep 5. Mol Aspects Med. 2015. PMID: 26344371 Review.
-
Laforin prevents stress-induced polyglucosan body formation and Lafora disease progression in neurons.Mol Neurobiol. 2013 Aug;48(1):49-61. doi: 10.1007/s12035-013-8438-2. Epub 2013 Apr 2. Mol Neurobiol. 2013. PMID: 23546741 Free PMC article.
-
Neurodegeneration and functional impairments associated with glycogen synthase accumulation in a mouse model of Lafora disease.EMBO Mol Med. 2011 Nov;3(11):667-81. doi: 10.1002/emmm.201100174. Epub 2011 Aug 29. EMBO Mol Med. 2011. PMID: 21882344 Free PMC article.
-
Astrocytes and neurons produce distinct types of polyglucosan bodies in Lafora disease.Glia. 2018 Oct;66(10):2094-2107. doi: 10.1002/glia.23463. Epub 2018 Aug 26. Glia. 2018. PMID: 30152044 Free PMC article.
Cited by
-
Wdfy3 regulates glycophagy, mitophagy, and synaptic plasticity.J Cereb Blood Flow Metab. 2021 Dec;41(12):3213-3231. doi: 10.1177/0271678X211027384. Epub 2021 Jun 29. J Cereb Blood Flow Metab. 2021. PMID: 34187232 Free PMC article.
-
Inactivation of Laforin Phosphatase and Increased Glucose Uptake Underlie Glycogen Synthase-Mediated Neuronal Survival Under Oxidative Stress.Mol Neurobiol. 2025 Sep;62(9):11145-11163. doi: 10.1007/s12035-025-04955-w. Epub 2025 Apr 22. Mol Neurobiol. 2025. PMID: 40261604
-
Dapagliflozin ameliorates Lafora disease phenotype in a zebrafish model.Biomed Pharmacother. 2025 Feb;183:117800. doi: 10.1016/j.biopha.2024.117800. Epub 2025 Jan 2. Biomed Pharmacother. 2025. PMID: 39753095 Free PMC article.
-
Brain glucose and ketone utilization in brain aging and neurodegenerative diseases.Int Rev Neurobiol. 2020;154:79-110. doi: 10.1016/bs.irn.2020.03.015. Epub 2020 Jul 10. Int Rev Neurobiol. 2020. PMID: 32739015 Free PMC article. Review.
-
Wnt5a Increases the Glycolytic Rate and the Activity of the Pentose Phosphate Pathway in Cortical Neurons.Neural Plast. 2016;2016:9839348. doi: 10.1155/2016/9839348. Epub 2016 Sep 5. Neural Plast. 2016. PMID: 27688915 Free PMC article.
References
-
- Akiyama H, Kameyama M, Akiguchi I, Sugiyama H, Kawamata T, Fukuyama H, Kimura H, Matsushita M, Takeda T. Periodic acid-Schiff (PAS)-positive, granular structures increase in the brain of senescence accelerated mouse (SAM) Acta Neuropathol. 1986;72:124–129. - PubMed
-
- Bouskila M, Hunter RW, Ibrahim AF, Delattre L, Peggie M, van Diepen JA, Voshol PJ, Jensen J, Sakamoto K. Allosteric regulation of glycogen synthase controls glycogen synthesis in muscle. Cell Metab. 2010;12:456–466. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Brown AM. Brain glycogen re-awakened. J. Neurochem. 2004;89:537–552. - PubMed
-
- Cataldo AM, Broadwell RD. Cytochemical identification of cerebral glycogen and glucose-6-phosphatase activity under normal and experimental conditions. II. Choroid plexus and ependymal epithelia, endothelia and pericytes. J. Neurocytol. 1986;15:511–524. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
