

Harnessing the unique structural properties of isolated α-helices
- PMID: 25059657
- PMCID: PMC4162150
- DOI: 10.1074/jbc.R114.583906
Harnessing the unique structural properties of isolated α-helices
Abstract
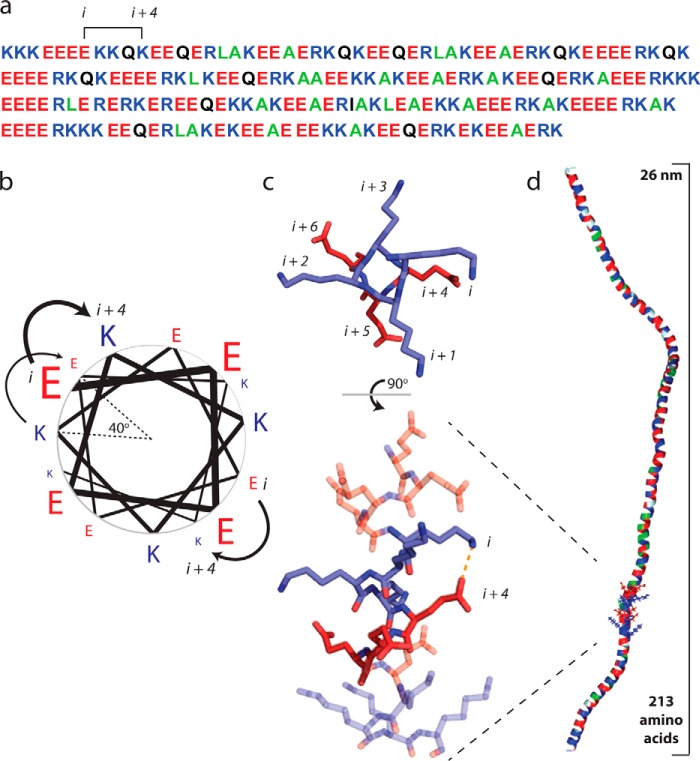
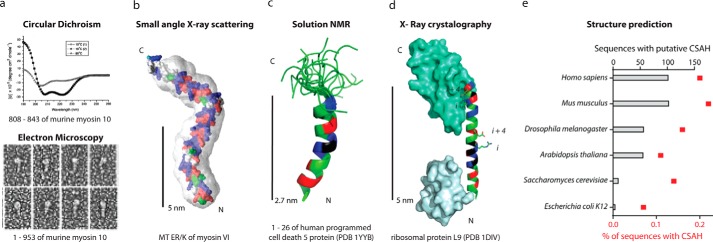
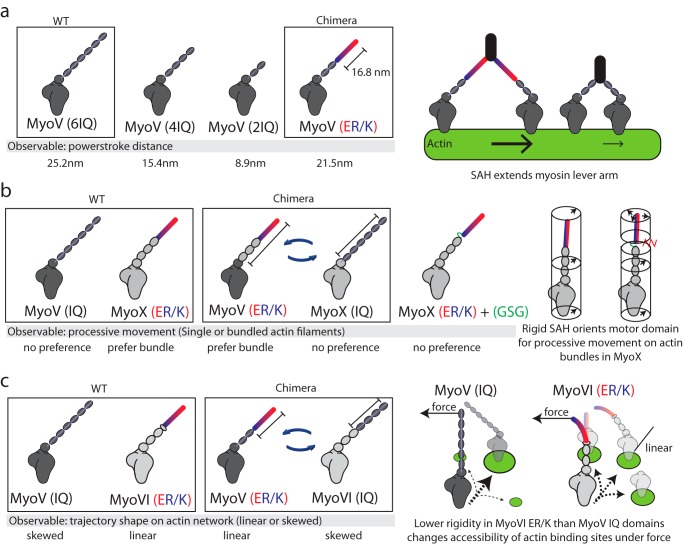
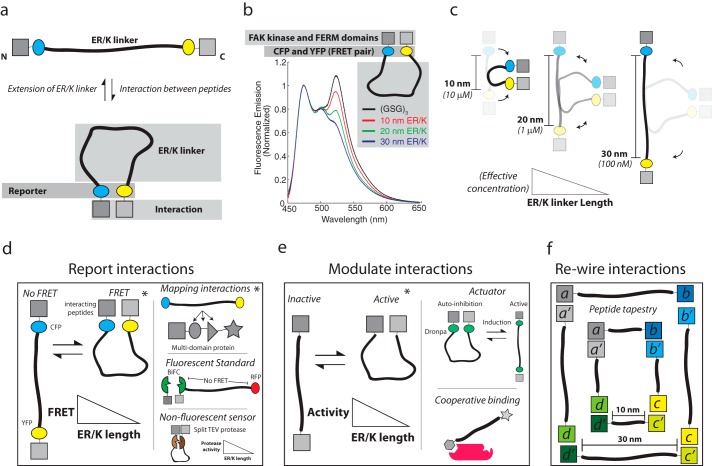
The α-helix is a ubiquitous secondary structural element that is almost exclusively observed in proteins when stabilized by tertiary or quaternary interactions. However, beginning with the unexpected observations of α-helix formation in the isolated C-peptide in ribonuclease A, there is growing evidence that a significant percentage (0.2%) of all proteins contain isolated stable single α-helical domains (SAH). These SAH domains provide unique structural features essential for normal protein function. A subset of SAH domains contain a characteristic ER/K motif, composed of a repeating sequence of ∼4 consecutive glutamic acids followed by ∼4 consecutive basic arginine or lysine (R/K) residues. The ER/K α-helix, also termed the ER/K linker, has been extensively characterized in the context of the myosin family of molecular motors and is emerging as a versatile structural element for protein and cellular engineering applications. Here, we review the structure and function of SAH domains, as well as the tools to identify them in natural proteins. We conclude with a discussion of recent studies that have successfully used the modular ER/K linker for engineering chimeric myosin proteins with altered mechanical properties, as well as synthetic polypeptides that can be used to monitor and systematically modulate protein interactions within cells.
Keywords: Biosensor; Coiled-coil; ER/K Alpha-Helix; ER/K Linker; Fluorescence Resonance Energy Transfer (FRET); Myosin; Protein Structure; Protein-Protein Interaction; Single Alpha-helical Domain (SAH).
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Knight P. J., Thirumurugan K., Xu Y., Wang F., Kalverda A. P., Stafford W. F., 3rd, Sellers J. R., Peckham M. (2005) The predicted coiled-coil domain of myosin 10 forms a novel elongated domain that lengthens the head. J. Biol. Chem. 280, 34702–34708 - PubMed
-
- Woolfson D. N. (2005) The design of coiled-coil structures and assemblies. Adv. Protein Chem. 70, 79–112 - PubMed
-
- Scholtz J. M., Baldwin R. L. (1992) The mechanism of α-helix formation by peptides. Annu. Rev. Biophys. Biomol. Struct. 21, 95–118 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
