

Sleep promotes cortical response potentiation following visual experience
- PMID: 25061244
- PMCID: PMC4098801
- DOI: 10.5665/sleep.3830
Sleep promotes cortical response potentiation following visual experience
Abstract
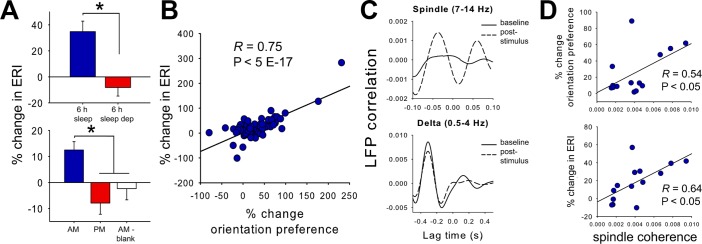
Study objectives: Sleep has been hypothesized to globally reduce synaptic strength. However, recent findings suggest that in the context of learning and memory consolidation, sleep may promote synaptic potentiation. We tested the requirement for sleep in a naturally occurring form of experience-dependent synaptic potentiation in the adult mouse visual cortex (V1), which is initiated by patterned visual experience.
Design: Visual responses were recorded in individual V1 neurons before and after presentation of an oriented grating stimulus, and after subsequent sleep or sleep deprivation.
Measurements and results: We find that V1 response potentiation-associated with a shift in orientation preference in favor of the presented stimulus-occurs only after sleep and only during the entrained circadian sleep phase, and is blocked by sleep deprivation. Induction of plasticity following stimulus presentation is associated with an increase in principal neuron firing in V1, which is present in all behavioral states and occurs regardless of time of day. Sleep dependent potentiation is proportional to phase-locking of neuronal activity with thalamocortical spindle oscillations.
Conclusions: Our results suggest that sleep can promote cortical synaptic potentiation in vivo, and that this potentiation may be mediated by slow wave sleep spindles.
Citation: Aton SJ, Suresh A, Broussard C, Frank MG. Sleep promotes cortical response potentiation following visual experience.
Keywords: in vivo recording; sleep; synaptic plasticity; thalamocortical oscillations; visual system.
Figures
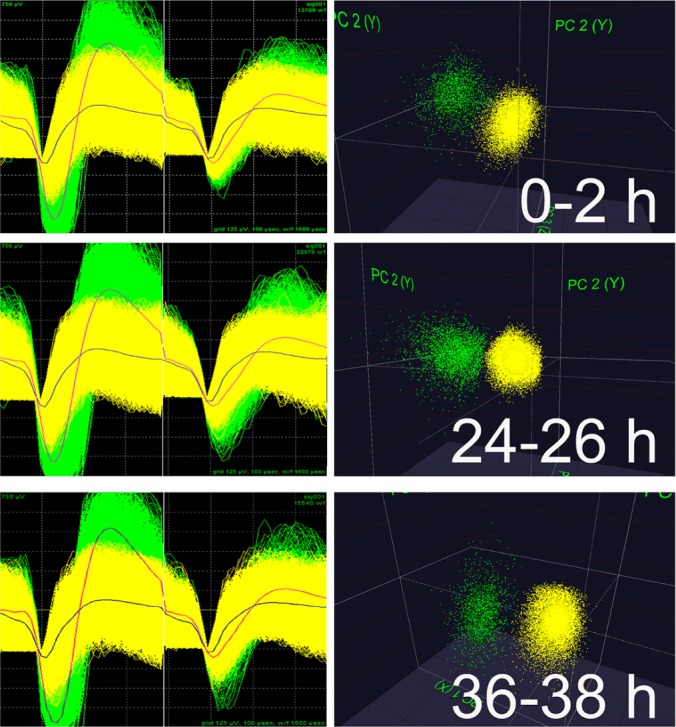
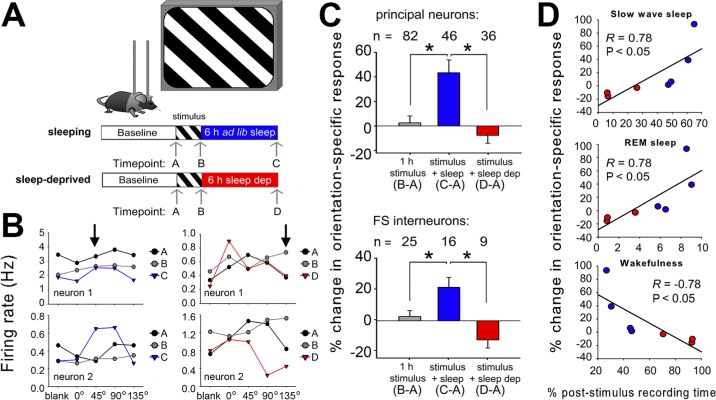
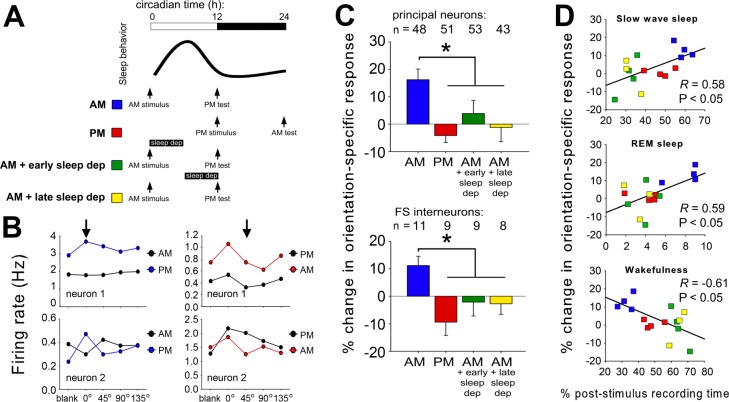
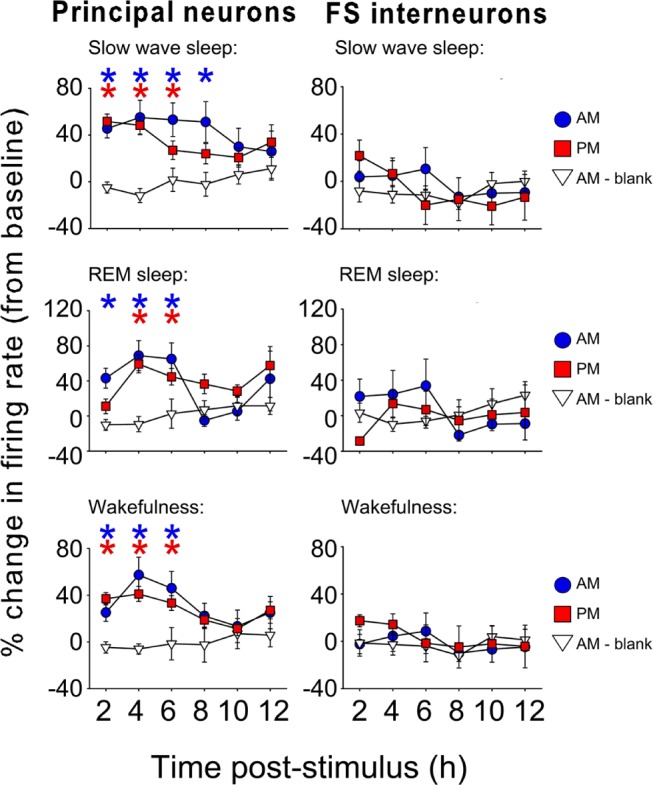
Comment in
- Sleep. 37:1157.
References
-
- Aton SJ, Seibt J, Frank MG. Encyclopedia of life science. Chichester: John Wiley and Sons, Ltd.; 2009. Sleep and memory.
-
- Tononi G, Cirelli C. Sleep and synaptic homeostasis: a hypothesis. Brain Res Bull. 2003;62:143–50. - PubMed
-
- Cirelli C, Gutierrez CM, Tononi G. Extensive and divergent effects of sleep and wakefulness on brain gene expression. Neuron. 2004;41:35–43. - PubMed
-
- Vyazovskiy VV, Cirelli C, Pfister-Genskow M, Faraguna U, Tononi G. Molecular and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep. 2009;63:865–78. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
