

Characterization of the small exported Plasmodium falciparum membrane protein SEMP1
- PMID: 25062022
- PMCID: PMC4111544
- DOI: 10.1371/journal.pone.0103272
Characterization of the small exported Plasmodium falciparum membrane protein SEMP1
Abstract
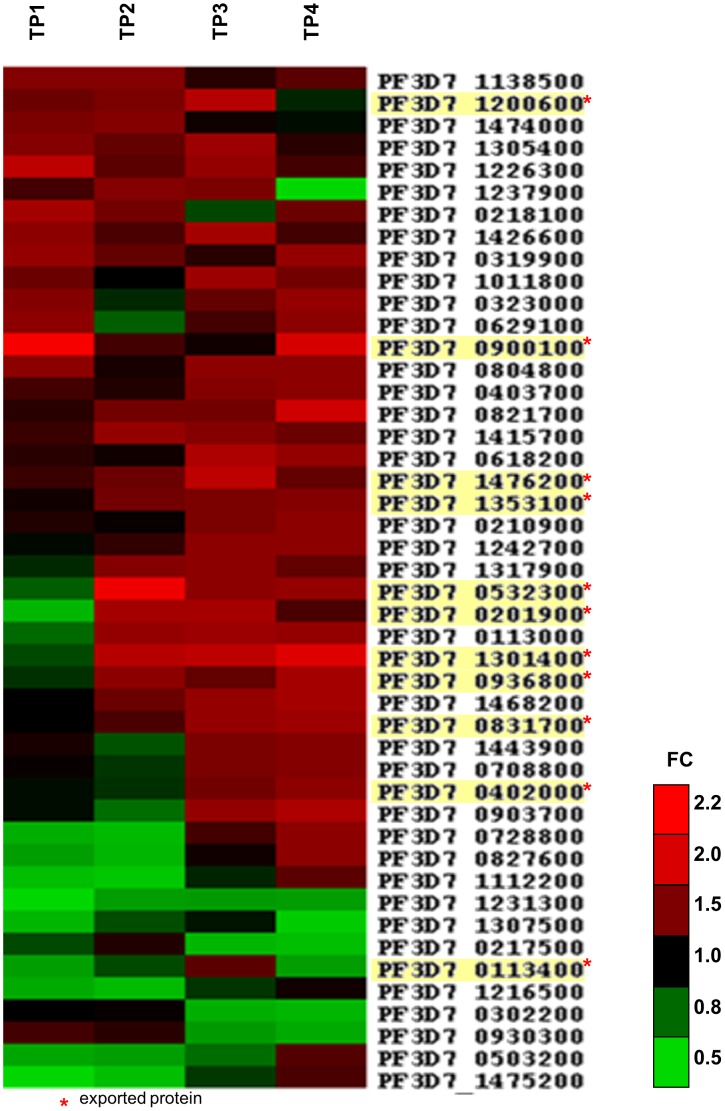
Survival and virulence of the human malaria parasite Plasmodium falciparum during the blood stage of infection critically depend on extensive host cell refurbishments mediated through export of numerous parasite proteins into the host cell. The parasite-derived membranous structures called Maurer's clefts (MC) play an important role in protein trafficking from the parasite to the red blood cell membrane. However, their specific function has yet to be determined. We identified and characterized a new MC membrane protein, termed small exported membrane protein 1 (SEMP1). Upon invasion it is exported into the RBC cytosol where it inserts into the MCs before it is partly translocated to the RBC membrane. Using conventional and conditional loss-of-function approaches we showed that SEMP1 is not essential for parasite survival, gametocytogenesis, or PfEMP1 export under culture conditions. Co-IP experiments identified several potential interaction partners, including REX1 and other membrane-associated proteins that were confirmed to co-localize with SEMP1 at MCs. Transcriptome analysis further showed that expression of a number of exported parasite proteins was up-regulated in SEMP1-depleted parasites. By using Co-IP and transcriptome analysis for functional characterization of an exported parasite protein we provide a new starting point for further detailed dissection and characterisation of MC-associated protein complexes.
Conflict of interest statement
Figures
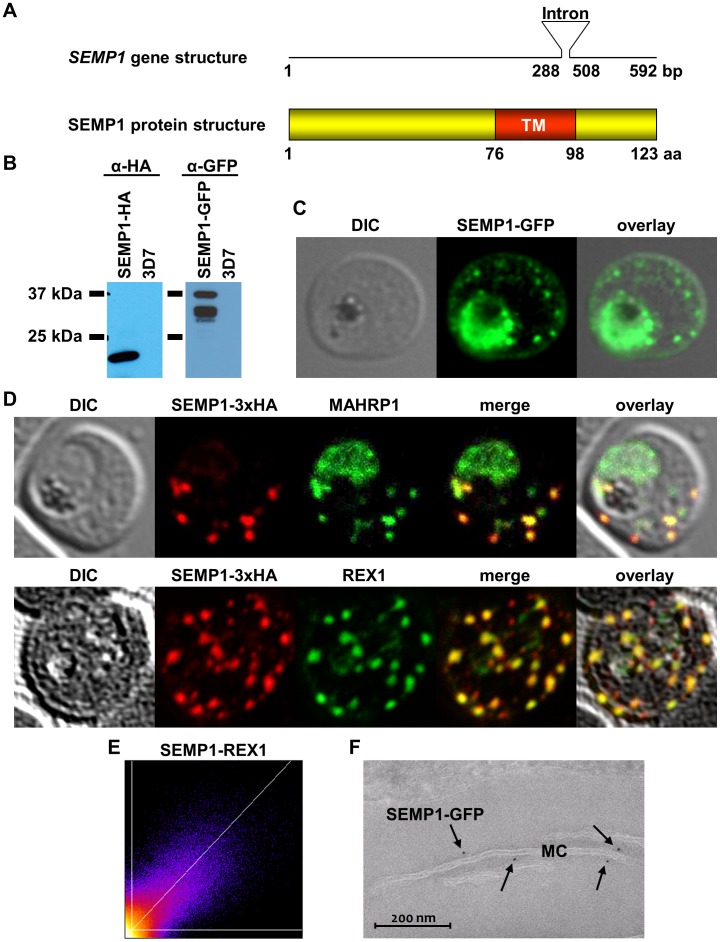
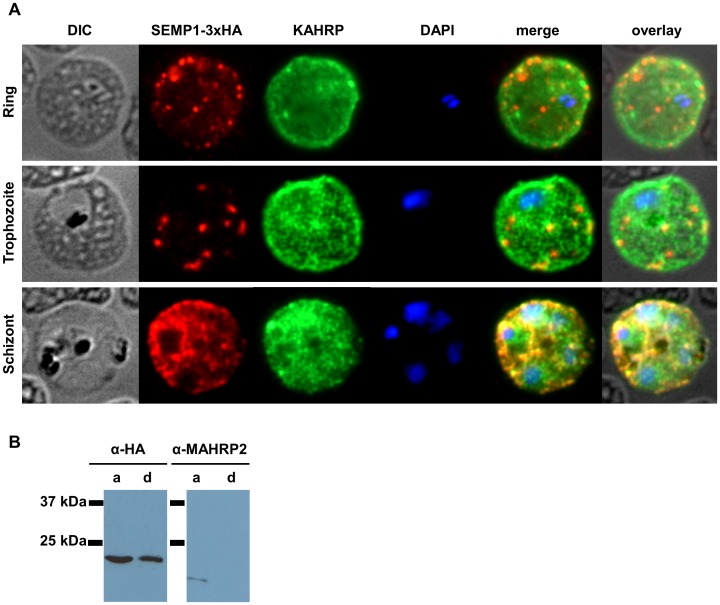
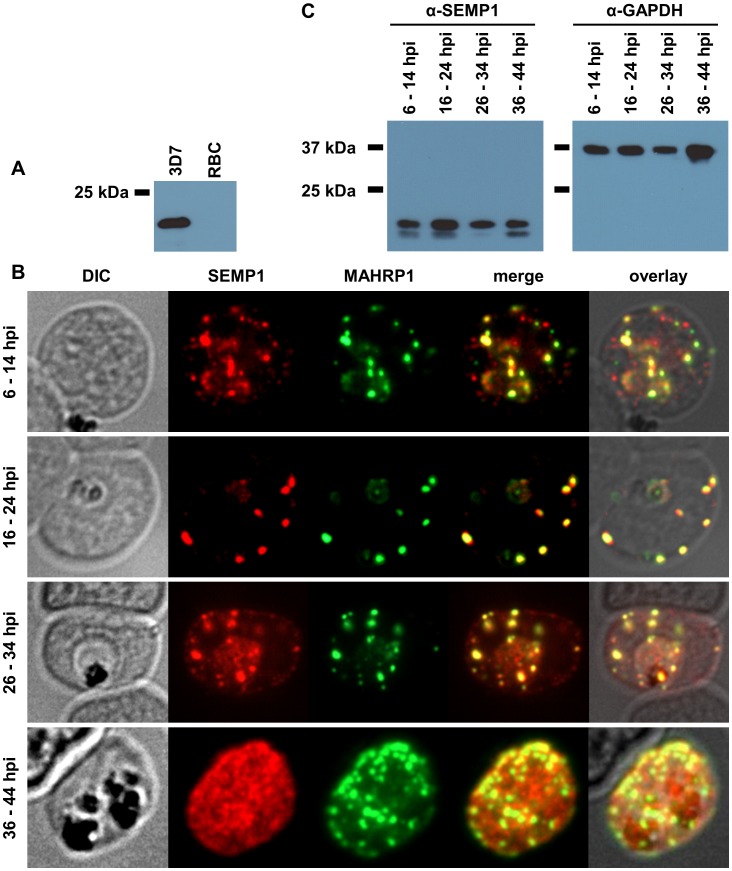
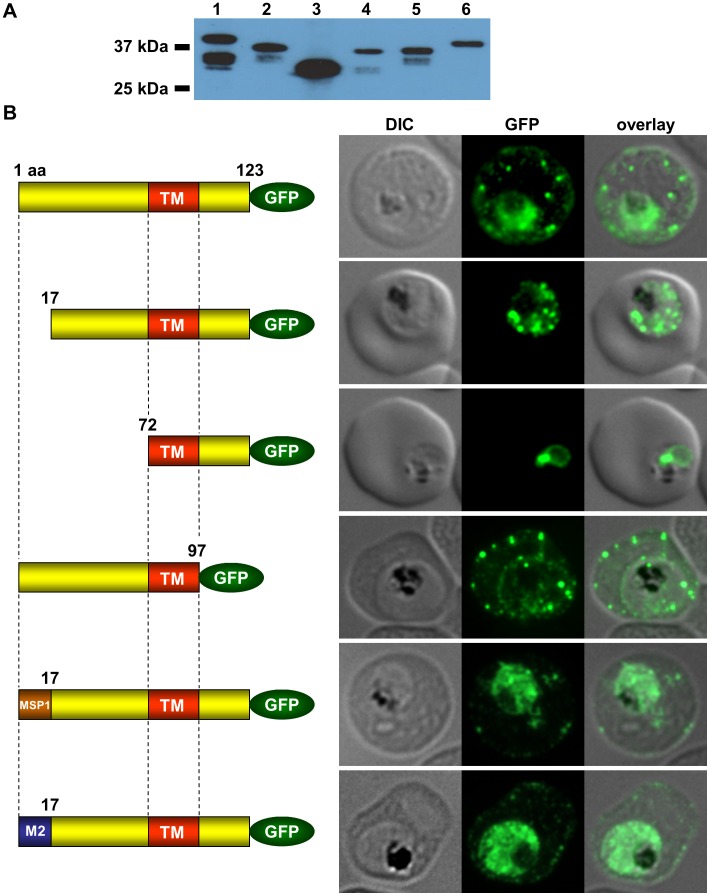
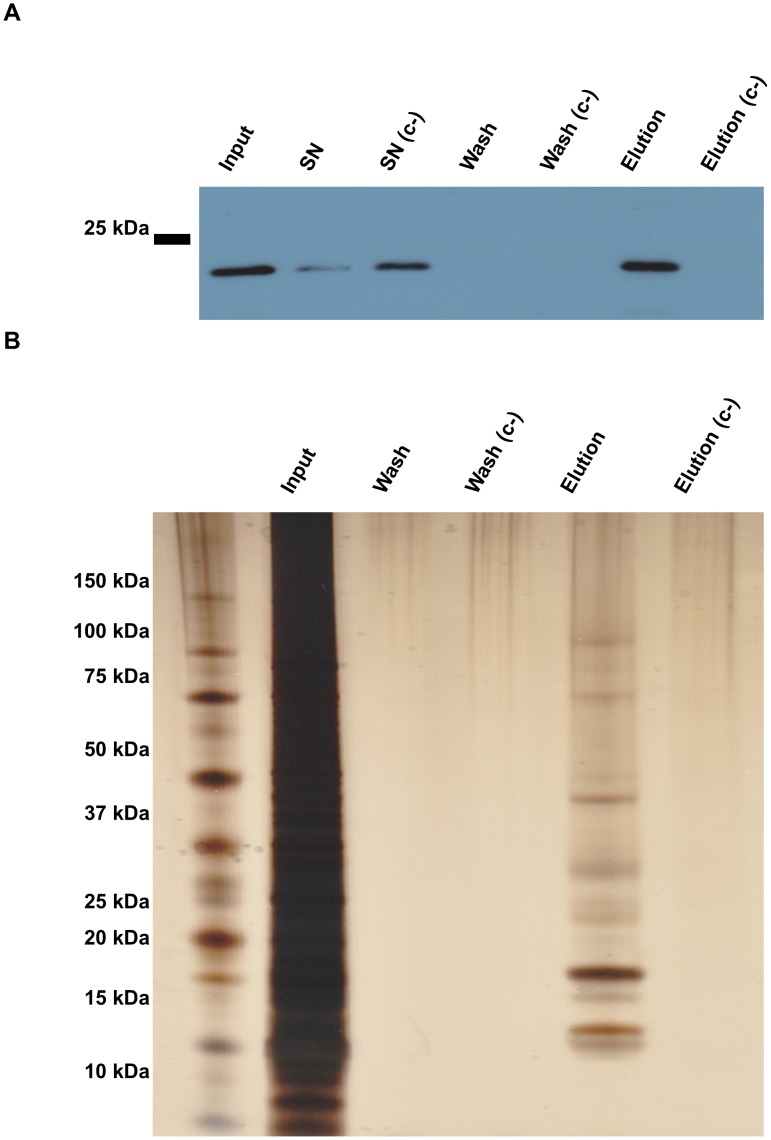
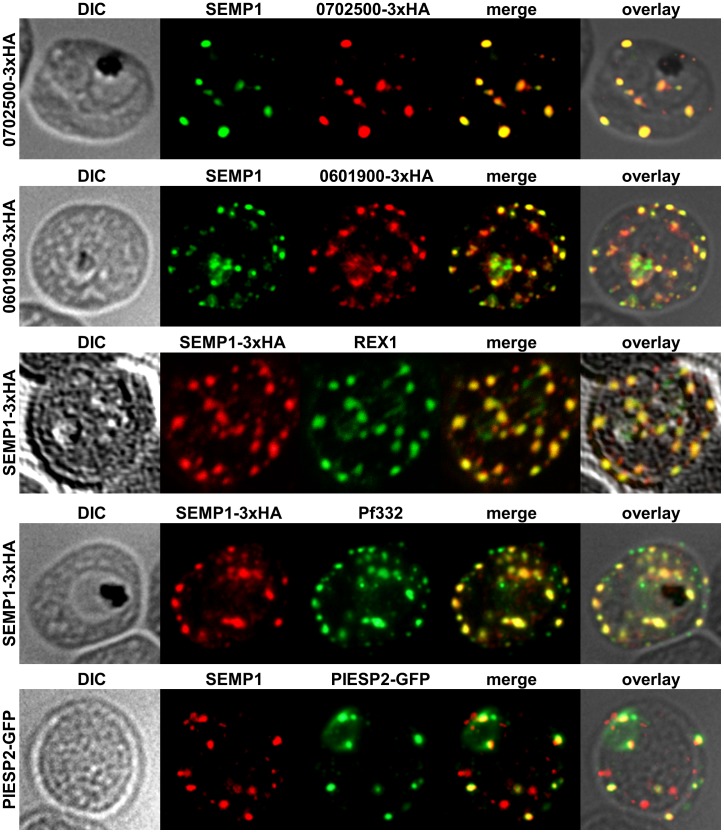
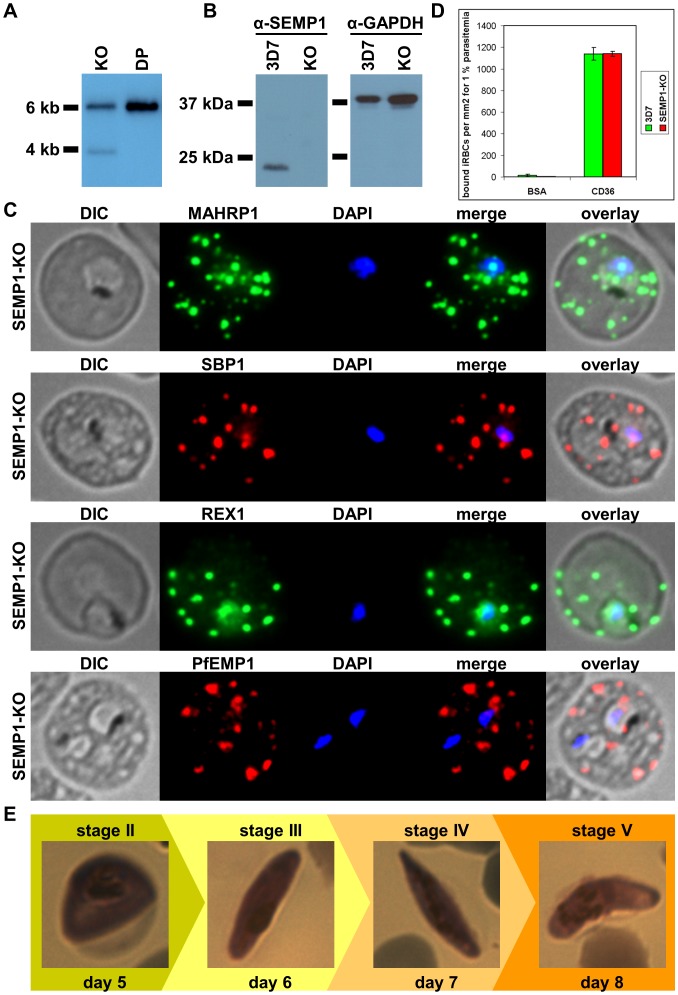
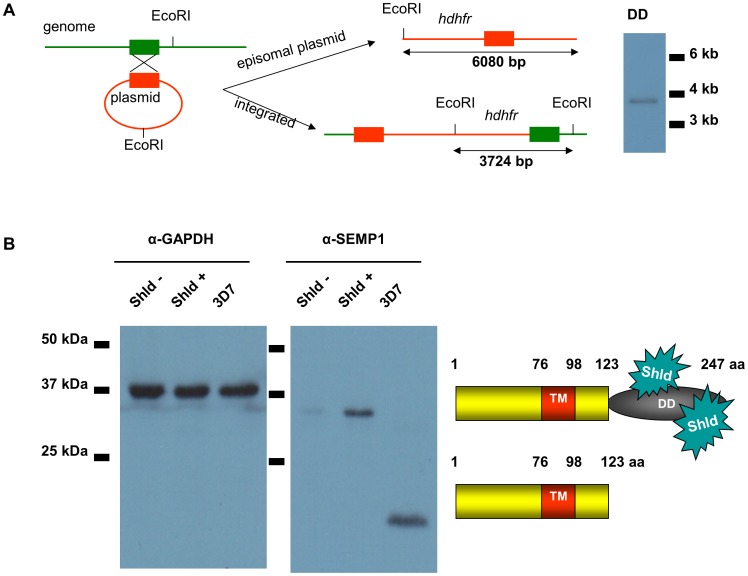
References
-
- World Health Organization (2012) WHO, World Health Report 2012.
-
- Miller LH, Baruch DI, Marsh K, Doumbo OK (2002) The pathogenic basis of malaria. Nature 415: 673–679. - PubMed
-
- Przyborski JM, Wickert H, Krohne G, Lanzer M (2003) Maurer's clefts—a novel secretory organelle? Mol. Biochem. Parasitol. 132: 17–26. - PubMed
-
- Hiller NL, Bhattacharjee S, van Ooij C, Liolios K, Harrison T, et al. (2004) A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science 306: 1934–1937. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
