

The DNA sensor, cyclic GMP-AMP synthase, is essential for induction of IFN-β during Chlamydia trachomatis infection
- PMID: 25070851
- PMCID: PMC4212656
- DOI: 10.4049/jimmunol.1302718
The DNA sensor, cyclic GMP-AMP synthase, is essential for induction of IFN-β during Chlamydia trachomatis infection
Abstract
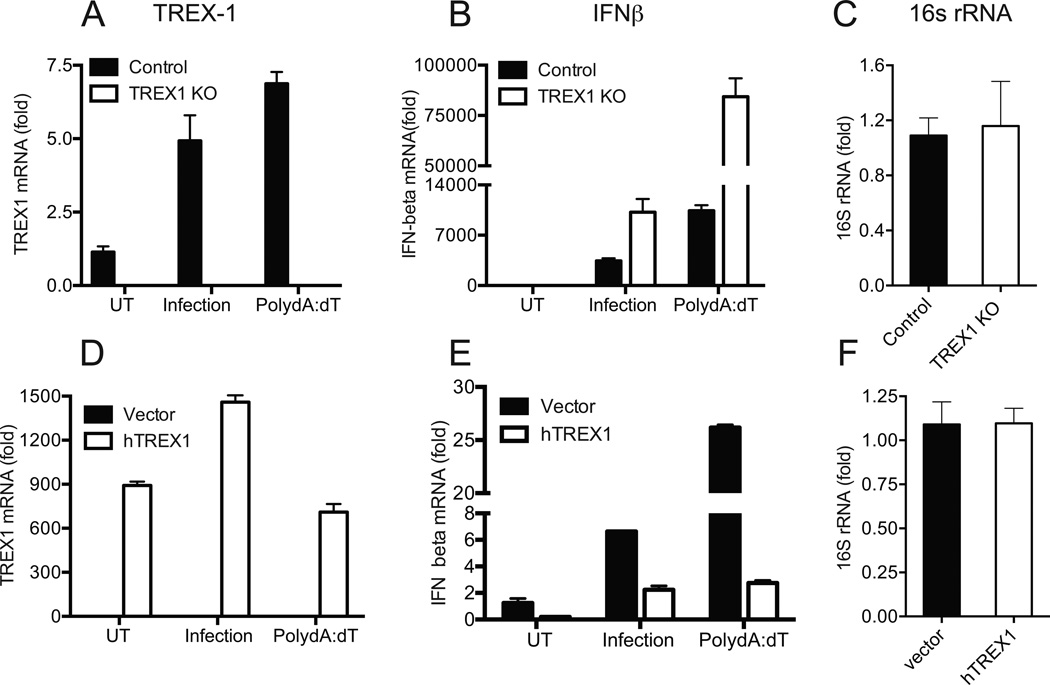
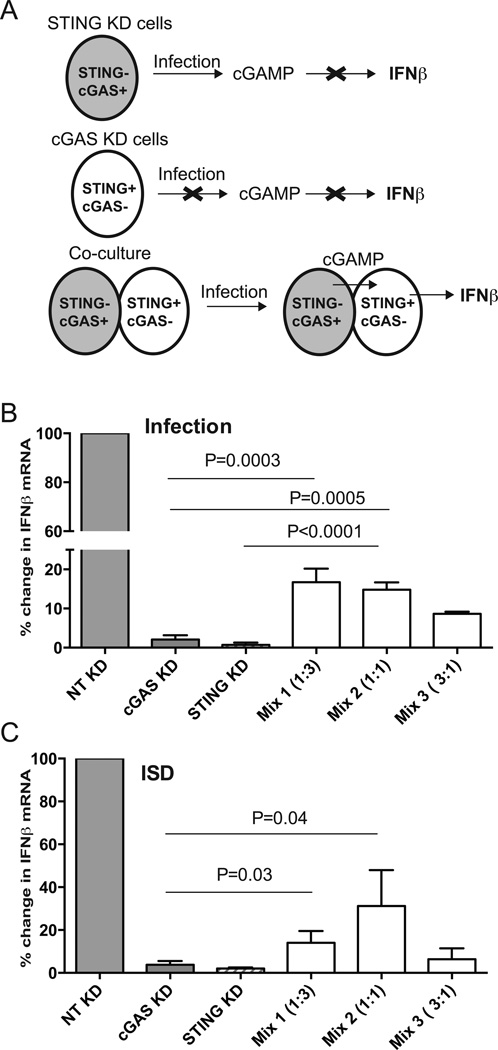
IFN-β has been implicated as an effector of oviduct pathology resulting from genital chlamydial infection in the mouse model. In this study, we investigated the role of cytosolic DNA and engagement of DNA sensors in IFN-β expression during chlamydial infection. We determined that three-prime repair exonuclease-1, a host 3' to 5' exonuclease, reduced IFN-β expression significantly during chlamydial infection using small interfering RNA and gene knockout fibroblasts, implicating cytosolic DNA as a ligand for this response. The DNA sensor cyclic GMP-AMP synthase (cGAS) has been shown to bind cytosolic DNA to generate cyclic GMP-AMP, which binds to the signaling adaptor stimulator of IFN genes (STING) to induce IFN-β expression. We determined that cGAS is required for IFN-β expression during chlamydial infection in multiple cell types. Interestingly, although infected cells deficient for STING or cGAS alone failed to induce IFN-β, coculture of cells depleted for either STING or cGAS rescued IFN-β expression. These data demonstrate that cyclic GMP-AMP produced in infected cGAS(+)STING(-) cells can migrate into adjacent cells via gap junctions to function in trans in cGAS(-)STING(+) cells. Furthermore, we observed cGAS localized in punctate regions on the cytosolic side of the chlamydial inclusion membrane in association with STING, indicating that chlamydial DNA is most likely recognized outside the inclusion as infection progresses. These novel findings provide evidence that cGAS-mediated DNA sensing directs IFN-β expression during Chlamydia trachomatis infection and suggest that effectors from infected cells can directly upregulate IFN-β expression in adjacent uninfected cells during in vivo infection, contributing to pathogenesis.
Copyright © 2014 by The American Association of Immunologists, Inc.
Figures






References
-
- Xia M, Bumgarner RE, Lampe MF, Stamm WE. Chlamydia trachomatis infection alters host cell transcription in diverse cellular pathways. J Infect Dis. 2003;187:424–434. - PubMed
-
- Rasmussen SJ, Eckmann L, Quayle AJ, Shen L, Zhang YX, Anderson DJ, Fierer J, Stephens RS, Kagnoff MF. Secretion of proinflammatory cytokines by epithelial cells in response to Chlamydia infection suggests a central role for epithelial cells in chlamydial pathogenesis. J Clin Invest. 1997;99:77–87. - PMC - PubMed
-
- Lad SP, Fukuda EY, Li J, de la Maza LM, Li E. Up-regulation of the JAK/STAT1 signal pathway during Chlamydia trachomatis infection. J Immunol. 2005;174:7186–7193. - PubMed
-
- Qiu H, Fan Y, Joyee AG, Wang S, Han X, Bai H, Jiao L, Rooijen NVan, Yang X. Type I IFNs enhance susceptibility to Chlamydia muridarum lung infection by enhancing apoptosis of local macrophages. J Immunol. 2008;181:2092–2102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
