

Neuroimaging, cognition, light and circadian rhythms
- PMID: 25071478
- PMCID: PMC4086398
- DOI: 10.3389/fnsys.2014.00126
Neuroimaging, cognition, light and circadian rhythms
Abstract
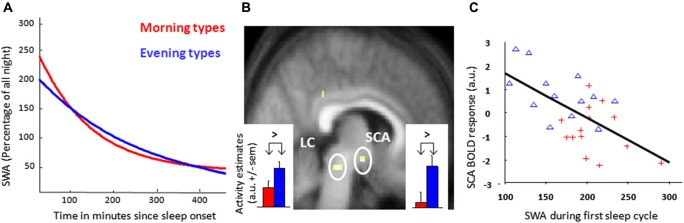
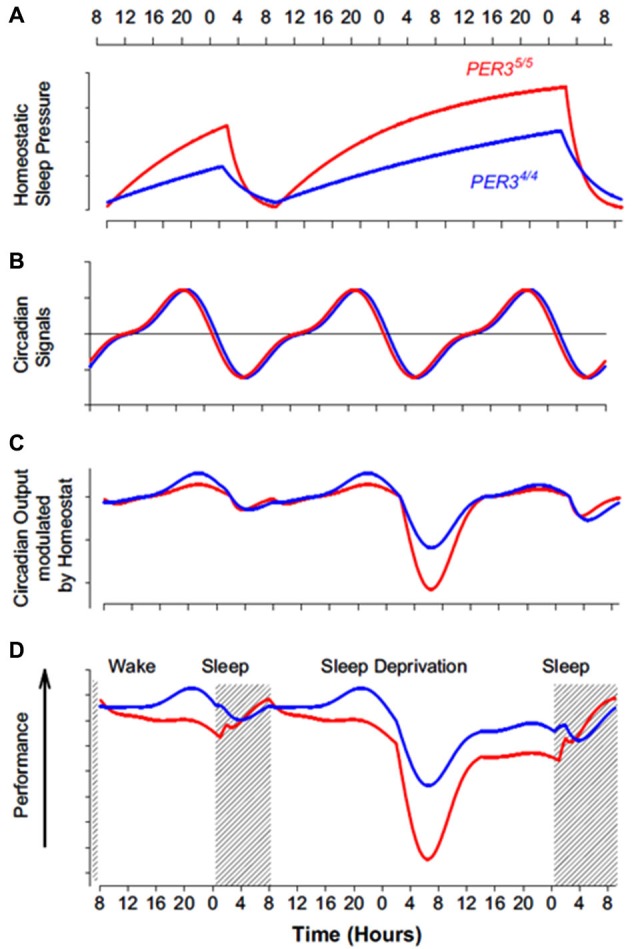
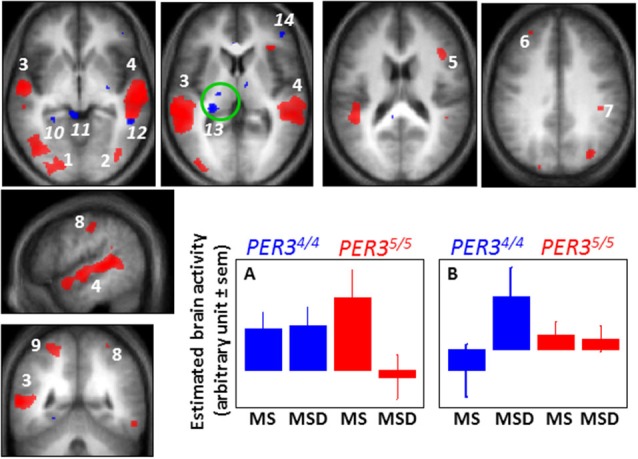
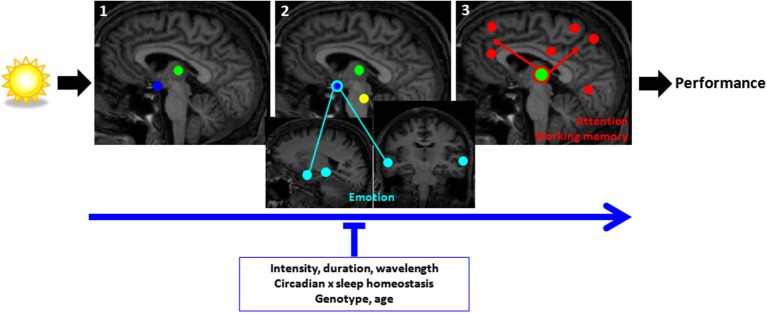
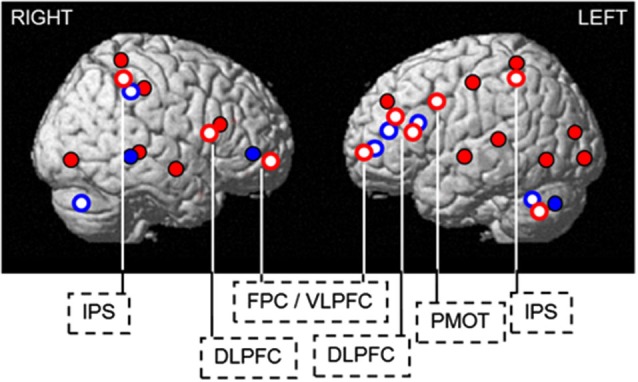
In humans, sleep and wakefulness and the associated cognitive processes are regulated through interactions between sleep homeostasis and the circadian system. Chronic disruption of sleep and circadian rhythmicity is common in our society and there is a need for a better understanding of the brain mechanisms regulating sleep, wakefulness and associated cognitive processes. This review summarizes recent investigations which provide first neural correlates of the combined influence of sleep homeostasis and circadian rhythmicity on cognitive brain activity. Markers of interindividual variations in sleep-wake regulation, such as chronotype and polymorphisms in sleep and clock genes, are associated with changes in cognitive brain responses in subcortical and cortical areas in response to manipulations of the sleep-wake cycle. This review also includes recent data showing that cognitive brain activity is regulated by light, which is a powerful modulator of cognition and alertness and also directly impacts sleep and circadian rhythmicity. The effect of light varied with age, psychiatric status, PERIOD3 genotype and changes in sleep homeostasis and circadian phase. These data provide new insights into the contribution of demographic characteristics, the sleep-wake cycle, circadian rhythmicity and light to brain functioning.
Keywords: circadian; cognition; fMRI; inter-individual differences in sleep-wake regulation; light; non-image-forming; non-visual; sleep.
Figures
References
-
- Archer S. N., Robilliard D. L., Skene D. J., Smits M., Williams A., Arendt J., et al. (2003). A length polymorphism in the circadian clock gene Per3 is linked to delayed sleep phase syndrome and extreme diurnal preference. Sleep 26, 413–415 - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources