

EGFR modulates DNA synthesis and repair through Tyr phosphorylation of histone H4
- PMID: 25073158
- PMCID: PMC4149291
- DOI: 10.1016/j.devcel.2014.06.008
EGFR modulates DNA synthesis and repair through Tyr phosphorylation of histone H4
Abstract
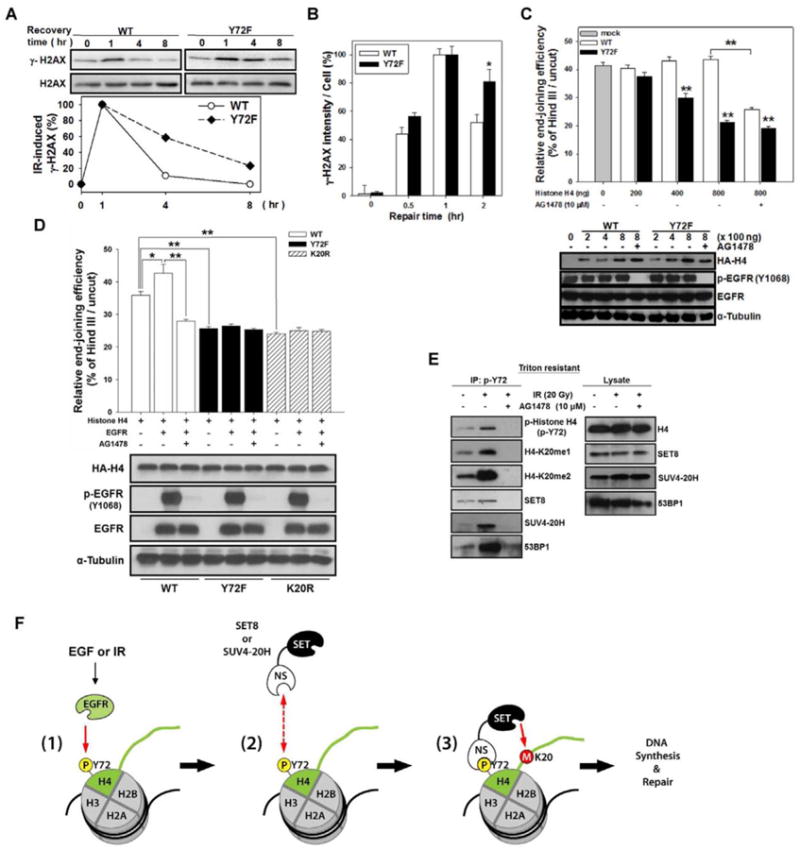
Posttranslational modifications of histones play fundamental roles in many biological functions. Specifically, histone H4-K20 methylation is critical for DNA synthesis and repair. However, little is known about how these functions are regulated by the upstream stimuli. Here, we identify a tyrosine phosphorylation site at Y72 of histone H4, which facilitates recruitment of histone methyltransferases (HMTases), SET8 and SUV4-20H, to enhance its K20 methylation, thereby promoting DNA synthesis and repair. Phosphorylation-defective histone H4 mutant is deficient in K20 methylation, leading to reduced DNA synthesis, delayed cell cycle progression, and decreased DNA repair ability. Disrupting the interaction between epidermal growth factor receptor (EGFR) and histone H4 by Y72 peptide significantly reduced tumor growth. Furthermore, EGFR expression clinically correlates with histone H4-Y72 phosphorylation, H4-K20 monomethylation, and the Ki-67 proliferation marker. These findings uncover a mechanism by which EGFR transduces signal to chromatin to regulate DNA synthesis and repair.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures






Similar articles
-
The SUV4-20H Histone Methyltransferases in Health and Disease.Int J Mol Sci. 2022 Apr 25;23(9):4736. doi: 10.3390/ijms23094736. Int J Mol Sci. 2022. PMID: 35563127 Free PMC article. Review.
-
Purification and functional characterization of SET8, a nucleosomal histone H4-lysine 20-specific methyltransferase.Curr Biol. 2002 Jul 9;12(13):1086-99. doi: 10.1016/s0960-9822(02)00924-7. Curr Biol. 2002. PMID: 12121615
-
Free and chromatin-associated mono-, di-, and trimethylation of histone H4-lysine 20 during development and cell cycle progression.Dev Biol. 2007 Apr 1;304(1):46-52. doi: 10.1016/j.ydbio.2006.12.019. Epub 2006 Dec 13. Dev Biol. 2007. PMID: 17229421 Free PMC article.
-
Certain and progressive methylation of histone H4 at lysine 20 during the cell cycle.Mol Cell Biol. 2008 Jan;28(1):468-86. doi: 10.1128/MCB.01517-07. Epub 2007 Oct 29. Mol Cell Biol. 2008. PMID: 17967882 Free PMC article.
-
Histone H4 lysine 20 methylation: key player in epigenetic regulation of genomic integrity.Nucleic Acids Res. 2013 Mar 1;41(5):2797-806. doi: 10.1093/nar/gkt012. Epub 2013 Jan 23. Nucleic Acids Res. 2013. PMID: 23345616 Free PMC article. Review.
Cited by
-
The SUV4-20H Histone Methyltransferases in Health and Disease.Int J Mol Sci. 2022 Apr 25;23(9):4736. doi: 10.3390/ijms23094736. Int J Mol Sci. 2022. PMID: 35563127 Free PMC article. Review.
-
Dynamic gene regulation by nuclear colony-stimulating factor 1 receptor in human monocytes and macrophages.Nat Commun. 2019 Apr 26;10(1):1935. doi: 10.1038/s41467-019-09970-9. Nat Commun. 2019. PMID: 31028249 Free PMC article.
-
Tyrosine kinase inhibitors protect the salivary gland from radiation damage by increasing DNA double-strand break repair.J Biol Chem. 2021 Jan-Jun;296:100401. doi: 10.1016/j.jbc.2021.100401. Epub 2021 Feb 9. J Biol Chem. 2021. PMID: 33571522 Free PMC article.
-
Cetuximab-induced natural killer cell cytotoxicity in head and neck squamous cell carcinoma cell lines: investigation of the role of cetuximab sensitivity and HPV status.Br J Cancer. 2020 Sep;123(5):752-761. doi: 10.1038/s41416-020-0934-3. Epub 2020 Jun 16. Br J Cancer. 2020. PMID: 32541873 Free PMC article.
-
A novel ligand-receptor relationship between families of ribonucleases and receptor tyrosine kinases.J Biomed Sci. 2018 Nov 19;25(1):83. doi: 10.1186/s12929-018-0484-7. J Biomed Sci. 2018. PMID: 30449278 Free PMC article. Review.
References
-
- Blagoev B, Kratchmarova I, Ong SE, Nielsen M, Foster LJ, Mann M. A proteomics strategy to elucidate functional protein-protein interactions applied to EGF signaling. Nat Biotechnol. 2003;21:315–318. - PubMed
-
- Chen HC, Appeddu PA, Isoda H, Guan JL. Phosphorylation of tyrosine 397 in focal adhesion kinase is required for binding phosphatidylinositol 3-kinase. J Biol Chem. 1996;271:26329–26334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
