

Antigen export during liver infection of the malaria parasite augments protective immunity
- PMID: 25073641
- PMCID: PMC4128355
- DOI: 10.1128/mBio.01321-14
Antigen export during liver infection of the malaria parasite augments protective immunity
Abstract
Protective immunity against preerythrocytic malaria parasite infection is difficult to achieve. Intracellular Plasmodium parasites likely minimize antigen presentation by surface-expressed major histocompatibility complex class I (MHC-I) molecules on infected cells, yet they actively remodel their host cells by export of parasite factors. Whether exported liver-stage proteins constitute better candidates for MHC-I antigen presentation to CD8(+) T lymphocytes remains unknown. Here, we systematically characterized the contribution of protein export to the magnitude of antigen-specific T-cell responses against Plasmodium berghei liver-stage parasites in C57BL/6 mice. We generated transgenic sporozoites that secrete a truncated ovalbumin (OVA) surrogate antigen only in the presence of an amino-terminal protein export element. Immunization with live attenuated transgenic sporozoites revealed that antigen export was not critical for CD8(+) T-cell priming but enhanced CD8(+) T-cell proliferation in the liver. Upon transfer of antigen-specific CD8(+) T cells, liver-stage parasites secreting the target protein were eliminated more efficiently. We conclude that Plasmodium parasites strictly control protein export during liver infection to minimize immune recognition. Strategies that enhance the discharge of parasite proteins into infected hepatocytes could improve the efficacy of candidate preerythrocytic malaria vaccines. Importance: Vaccine development against Plasmodium parasites remains a priority in malaria research. The most advanced malaria subunit vaccine candidates contain Plasmodium surface proteins with important roles for parasite vital functions. A fundamental question is whether recognition by effector CD8(+) T cells is restricted to sporozoite surface antigens or extends to parasite proteins that are synthesized during the extensive parasite expansion phase in the liver. Using a surrogate model antigen, we found that a cytoplasmic antigen is able to induce robust protective CD8(+) T-cell responses, but protein export further enhances immunogenicity and protection. Our results show that a cytoplasmic localization does not exclude a protein's candidacy for malaria subunit vaccines and that protein secretion can enhance protective immunity.
Copyright © 2014 Montagna et al.
Figures
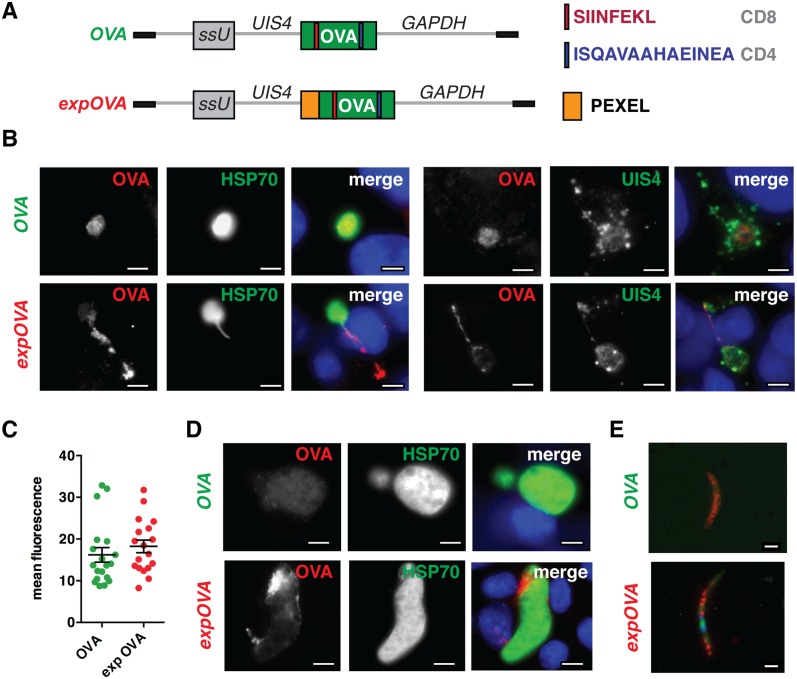
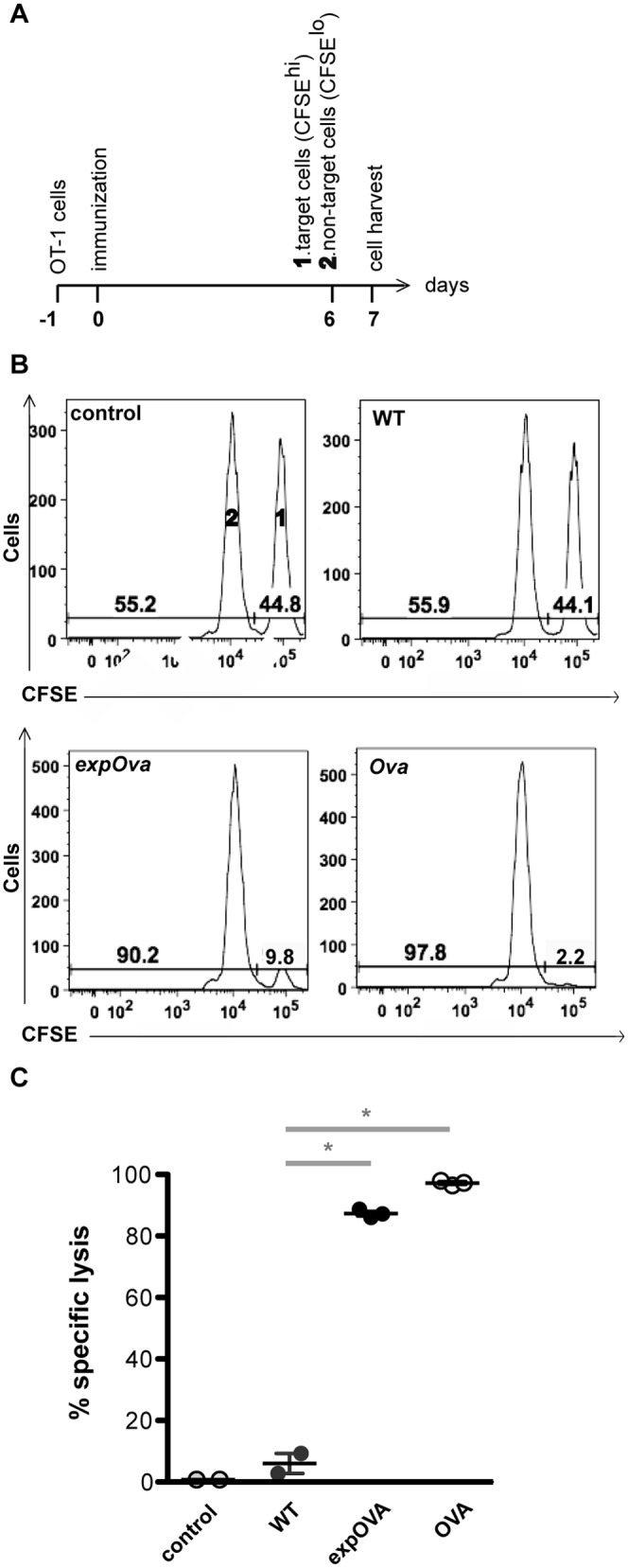

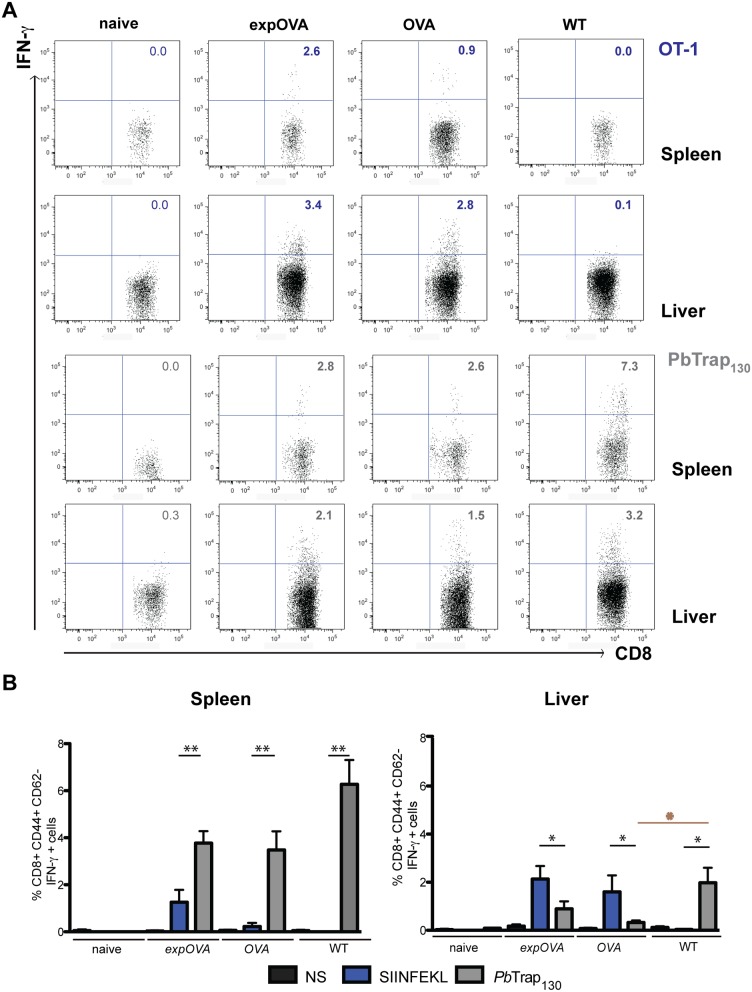
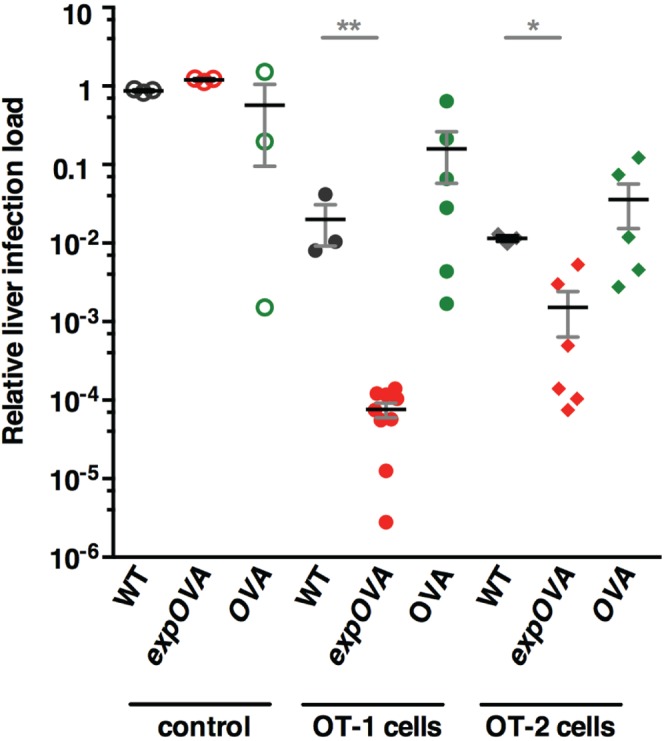
Similar articles
-
Discriminating Protective from Nonprotective Plasmodium-Specific CD8+ T Cell Responses.J Immunol. 2016 May 15;196(10):4253-62. doi: 10.4049/jimmunol.1600155. Epub 2016 Apr 15. J Immunol. 2016. PMID: 27084099 Free PMC article.
-
Importance of the Immunodominant CD8+ T Cell Epitope of Plasmodium berghei Circumsporozoite Protein in Parasite- and Vaccine-Induced Protection.Infect Immun. 2020 Sep 18;88(10):e00383-20. doi: 10.1128/IAI.00383-20. Print 2020 Sep 18. Infect Immun. 2020. PMID: 32719159 Free PMC article.
-
Direct processing and presentation of antigen from malaria sporozoites by professional antigen-presenting cells in the induction of CD8 T-cell responses.Immunol Cell Biol. 2005 Jun;83(3):307-12. doi: 10.1111/j.1440-1711.2005.01325.x. Immunol Cell Biol. 2005. PMID: 15877610
-
Protective CD8 T cells against Plasmodium liver stages: immunobiology of an 'unnatural' immune response.Immunol Rev. 2008 Oct;225:272-83. doi: 10.1111/j.1600-065X.2008.00671.x. Immunol Rev. 2008. PMID: 18837788 Free PMC article. Review.
-
Current Challenges in the Identification of Pre-Erythrocytic Malaria Vaccine Candidate Antigens.Front Immunol. 2020 Feb 21;11:190. doi: 10.3389/fimmu.2020.00190. eCollection 2020. Front Immunol. 2020. PMID: 32153565 Free PMC article. Review.
Cited by
-
Discriminating Protective from Nonprotective Plasmodium-Specific CD8+ T Cell Responses.J Immunol. 2016 May 15;196(10):4253-62. doi: 10.4049/jimmunol.1600155. Epub 2016 Apr 15. J Immunol. 2016. PMID: 27084099 Free PMC article.
-
Shedding of host autophagic proteins from the parasitophorous vacuolar membrane of Plasmodium berghei.Sci Rep. 2017 May 19;7(1):2191. doi: 10.1038/s41598-017-02156-7. Sci Rep. 2017. PMID: 28526861 Free PMC article.
-
The human immune system's response to carcinogenic and other infectious agents transmitted by mosquito vectors.Parasitol Res. 2017 Jan;116(1):1-9. doi: 10.1007/s00436-016-5272-2. Epub 2016 Oct 27. Parasitol Res. 2017. PMID: 27785601 Review.
-
A P. falciparum NF54 Reporter Line Expressing mCherry-Luciferase in Gametocytes, Sporozoites, and Liver-Stages.Front Cell Infect Microbiol. 2019 Apr 16;9:96. doi: 10.3389/fcimb.2019.00096. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31058097 Free PMC article.
-
The Plasmodium berghei translocon of exported proteins reveals spatiotemporal dynamics of tubular extensions.Sci Rep. 2015 Jul 29;5:12532. doi: 10.1038/srep12532. Sci Rep. 2015. PMID: 26219962 Free PMC article.
References
-
- Roestenberg M, McCall M, Hopman J, Wiersma J, Luty AJ, van Gemert GJ, van de Vegte-Bolmer M, van Schaijk B, Teelen K, Arens T, Spaarman L, de Mast Q, Roeffen W, Snounou G, Renia L, van der Ven A, Hermsen CC, Sauerwein R. 2009. Protection against a malaria challenge by sporozoite inoculation. N. Engl. J. Med. 361:468–477. 10.1056/NEJMoa0805832 - DOI - PubMed
-
- Roestenberg M, Teirlinck AC, McCall MB, Teelen K, Makamdop KN, Wiersma J, Arens T, Beckers P, van Gemert G, van de Vegte-Bolmer M, van der Ven AJ, Luty AJ, Hermsen CC, Sauerwein RW. 2011. Long-term protection against malaria after experimental sporozoite inoculation: an open-label follow-up study. Lancet 377:1770–1776. 10.1016/S0140-6736(11)60360-7 - DOI - PubMed
-
- Seder RA, Chang LJ, Enama ME, Zephir KL, Sarwar UN, Gordon IJ, Holman LA, James ER, Billingsley PF, Gunasekera A, Richman A, Chakravarty S, Manoj A, Velmurugan S, Li M, Ruben AJ, Li T, Eappen AG, Stafford RE, Plummer SH, Hendel CS, Novik L, Costner PJ, Mendoza FH, Saunders JG, Nason MC, Richardson JH, Murphy J, Davidson SA, Richie TL, Sedegah M, Sutamihardja A, Fahle GA, Lyke KE, Laurens MB, Roederer M, Tewari K, Epstein JE, Sim BK, Ledgerwood JE, Graham BS, Hoffman SL, VRC 312 Study Team 2013. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 341:1359–1365. 10.1126/science.1241800 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
