

Inactivation of thyA in Staphylococcus aureus attenuates virulence and has a strong impact on metabolism and virulence gene expression
- PMID: 25073642
- PMCID: PMC4128360
- DOI: 10.1128/mBio.01447-14
Inactivation of thyA in Staphylococcus aureus attenuates virulence and has a strong impact on metabolism and virulence gene expression
Abstract
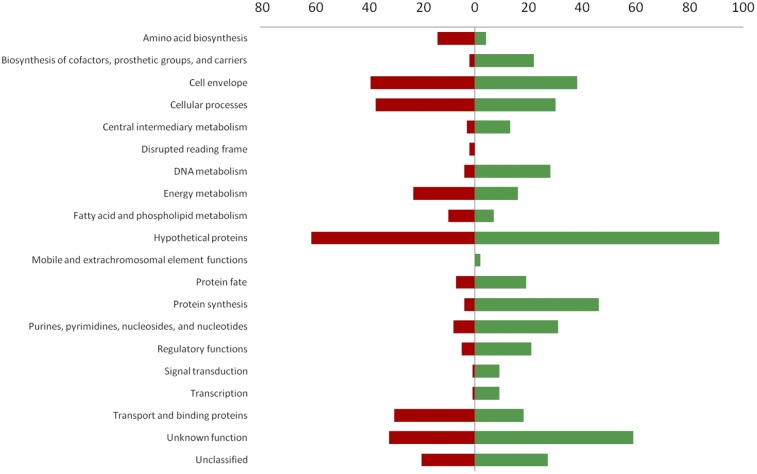
Staphylococcus aureus thymidine-dependent small-colony variants (TD-SCVs) are frequently isolated from patients with chronic S. aureus infections after long-term treatment with trimethoprim-sulfamethoxazole (TMP-SMX). While it has been shown that TD-SCVs were associated with mutations in thymidylate synthase (TS; thyA), the impact of such mutations on protein function is lacking. In this study, we showed that mutations in thyA were leading to inactivity of TS proteins, and TS inactivity led to tremendous impact on S. aureus physiology and virulence. Whole DNA microarray analysis of the constructed ΔthyA mutant identified severe alterations compared to the wild type. Important virulence regulators (agr, arlRS, sarA) and major virulence determinants (hla, hlb, sspAB, and geh) were downregulated, while genes important for colonization (fnbA, fnbB, spa, clfB, sdrC, and sdrD) were upregulated. The expression of genes involved in pyrimidine and purine metabolism and nucleotide interconversion changed significantly. NupC was identified as a major nucleoside transporter, which supported growth of the mutant during TMP-SMX exposure by uptake of extracellular thymidine. The ΔthyA mutant was strongly attenuated in virulence models, including a Caenorhabditis elegans killing model and an acute pneumonia mouse model. This study identified inactivation of TS as the molecular basis of clinical TD-SCV and showed that thyA activity has a major role for S. aureus virulence and physiology. Importance: Thymidine-dependent small-colony variants (TD-SCVs) of Staphylococcus aureus carry mutations in the thymidylate synthase (TS) gene (thyA) responsible for de novo synthesis of thymidylate, which is essential for DNA synthesis. TD-SCVs have been isolated from patients treated for long periods with trimethoprim-sulfamethoxazole (TMP-SMX) and are associated with chronic and recurrent infections. In the era of community-associated methicillin-resistant S. aureus, the therapeutic use of TMP-SMX is increasing. Today, the emergence of TD-SCVs is still underestimated due to misidentification in the diagnostic laboratory. This study showed for the first time that mutational inactivation of TS is the molecular basis for the TD-SCV phenotype and that TS inactivation has a strong impact on S. aureus virulence and physiology. Our study helps to understand the clinical nature of TD-SCVs, which emerge frequently once patients are treated with TMP-SMX.
Copyright © 2014 Kriegeskorte et al.
Figures







Similar articles
-
In vivo mutations of thymidylate synthase (encoded by thyA) are responsible for thymidine dependency in clinical small-colony variants of Staphylococcus aureus.J Bacteriol. 2008 Feb;190(3):834-42. doi: 10.1128/JB.00912-07. Epub 2007 Sep 28. J Bacteriol. 2008. PMID: 17905979 Free PMC article.
-
Thymidine-Dependent Staphylococcus aureus Small-Colony Variants Are Induced by Trimethoprim-Sulfamethoxazole (SXT) and Have Increased Fitness during SXT Challenge.Antimicrob Agents Chemother. 2015 Dec;59(12):7265-72. doi: 10.1128/AAC.00742-15. Epub 2015 Sep 14. Antimicrob Agents Chemother. 2015. PMID: 26369968 Free PMC article.
-
Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus.Int J Med Microbiol. 2007 Jul;297(4):217-25. doi: 10.1016/j.ijmm.2007.02.003. Epub 2007 Apr 6. Int J Med Microbiol. 2007. PMID: 17412637
-
Small colony variants of Staphylococcus aureus--review.Folia Microbiol (Praha). 2010 Nov;55(6):548-58. doi: 10.1007/s12223-010-0089-3. Epub 2011 Jan 21. Folia Microbiol (Praha). 2010. PMID: 21253898 Review.
-
Staphylococcus aureus Small Colony Variants (SCVs): a road map for the metabolic pathways involved in persistent infections.Front Cell Infect Microbiol. 2014 Jul 28;4:99. doi: 10.3389/fcimb.2014.00099. eCollection 2014. Front Cell Infect Microbiol. 2014. PMID: 25120957 Free PMC article. Review.
Cited by
-
Phenotypic and genotypic characterisation of thymine auxotrophy in Escherichia coli isolated from a patient with recurrent bloodstream infection.PLoS One. 2022 Jul 8;17(7):e0270256. doi: 10.1371/journal.pone.0270256. eCollection 2022. PLoS One. 2022. PMID: 35802671 Free PMC article.
-
Pyrimidine sufficiency is required for Sae two-component system signaling in Staphylococcus aureus.bioRxiv [Preprint]. 2025 Mar 20:2025.03.20.644390. doi: 10.1101/2025.03.20.644390. bioRxiv. 2025. Update in: J Bacteriol. 2025 Aug 21;207(8):e0011525. doi: 10.1128/jb.00115-25. PMID: 40166268 Free PMC article. Updated. Preprint.
-
The Electron Transport Chain Sensitizes Staphylococcus aureus and Enterococcus faecalis to the Oxidative Burst.Infect Immun. 2017 Nov 17;85(12):e00659-17. doi: 10.1128/IAI.00659-17. Print 2017 Dec. Infect Immun. 2017. PMID: 28993457 Free PMC article.
-
Functional diversity of staphylococcal surface proteins at the host-microbe interface.Front Microbiol. 2023 May 18;14:1196957. doi: 10.3389/fmicb.2023.1196957. eCollection 2023. Front Microbiol. 2023. PMID: 37275142 Free PMC article. Review.
-
Genome-wide analysis of fitness determinants of Staphylococcus aureus during growth in milk.PLoS Pathog. 2025 Apr 9;21(4):e1013080. doi: 10.1371/journal.ppat.1013080. eCollection 2025 Apr. PLoS Pathog. 2025. PMID: 40203072 Free PMC article.
References
-
- Vandenesch F, Naimi T, Enright MC, Lina G, Nimmo GR, Heffernan H, Liassine N, Bes M, Greenland T, Reverdy ME, Etienne J. 2003. Community-acquired methicillin-resistant Staphylococcus aureus carrying the Pantón-Valentine leukocidin genes: worldwide emergence. Emerg. Infect. Dis. 9:978–984. 10.3201/eid0908.030089 - DOI - PMC - PubMed
-
- Köck R, Brakensiek L, Mellmann A, Kipp F, Henderikx M, Harmsen D, Daniels-Haardt I, von Eiff C, Becker K, Hendrix MG, Friedrich AW. 2009. Cross-border comparison of the admission prevalence and clonal structure of methicillin-resistant Staphylococcus aureus. J. Hosp. Infect. 71:320–326. 10.1016/j.jhin.2008.12.001 - DOI - PubMed
-
- Proctor RA, Balwit JM, Vesga O. 1994. Variant subpopulations of Staphylococcus aureus as cause of persistent and recurrent infections. Infect. Agents Dis. 3:302–312 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
