

Copper-catalyzed azide-alkyne cycloaddition (click chemistry)-based detection of global pathogen-host AMPylation on self-assembled human protein microarrays
- PMID: 25073739
- PMCID: PMC4223499
- DOI: 10.1074/mcp.M114.041103
Copper-catalyzed azide-alkyne cycloaddition (click chemistry)-based detection of global pathogen-host AMPylation on self-assembled human protein microarrays
Abstract
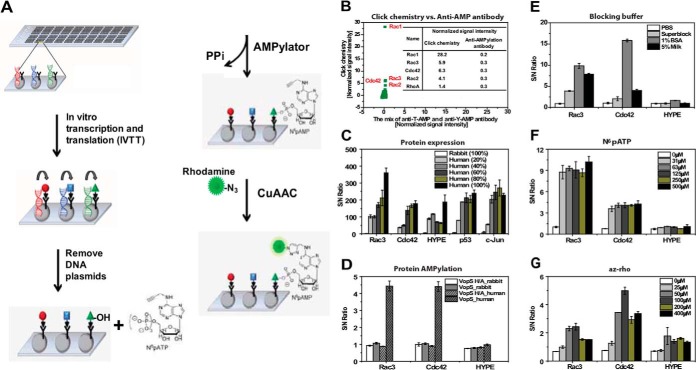
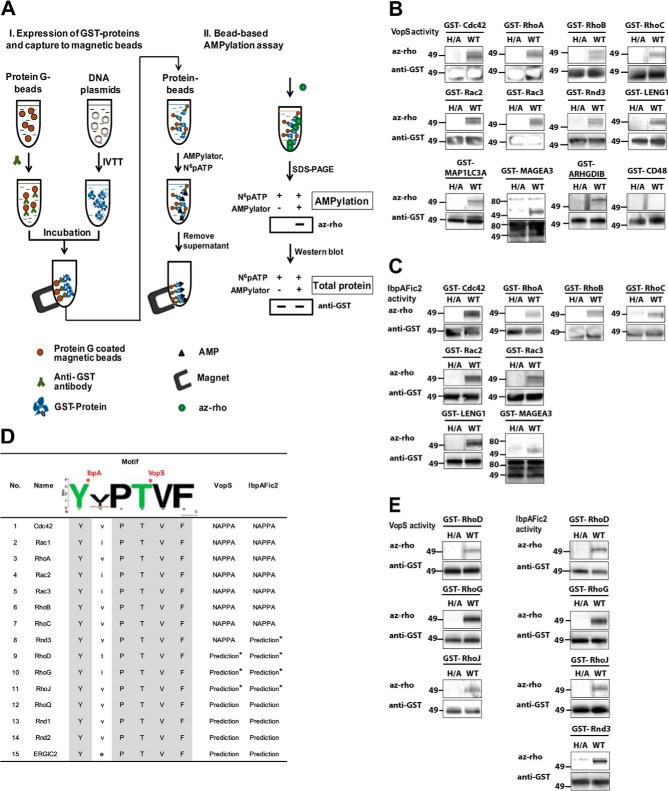
AMPylation (adenylylation) is a recently discovered mechanism employed by infectious bacteria to regulate host cell signaling. However, despite significant effort, only a few host targets have been identified, limiting our understanding of how these pathogens exploit this mechanism to control host cells. Accordingly, we developed a novel nonradioactive AMPylation screening platform using high-density cell-free protein microarrays displaying human proteins produced by human translational machinery. We screened 10,000 unique human proteins with Vibrio parahaemolyticus VopS and Histophilus somni IbpAFic2, and identified many new AMPylation substrates. Two of these, Rac2, and Rac3, were confirmed in vivo as bona fide substrates during infection with Vibrio parahaemolyticus. We also mapped the site of AMPylation of a non-GTPase substrate, LyGDI, to threonine 51, in a region regulated by Src kinase, and demonstrated that AMPylation prevented its phosphorylation by Src. Our results greatly expanded the repertoire of potential host substrates for bacterial AMPylators, determined their recognition motif, and revealed the first pathogen-host interaction AMPylation network. This approach can be extended to identify novel substrates of AMPylators with different domains or in different species and readily adapted for other post-translational modifications.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures





References
-
- Muller M. P., Peters H., Blumer J., Blankenfeldt W., Goody R. S., Itzen A. (2010) The Legionella effector protein DrrA AMPylates the membrane traffic regulator Rab1b. Science 329, 946–949 - PubMed
-
- Yarbrough M. L., Li Y., Kinch L. N., Grishin N. V., Ball H. L., Orth K. (2009) AMPylation of Rho GTPases by Vibrio VopS disrupts effector binding and downstream signaling. Science 323, 269–272 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
