

Rag defects and thymic stroma: lessons from animal models
- PMID: 25076946
- PMCID: PMC4114104
- DOI: 10.3389/fimmu.2014.00259
Rag defects and thymic stroma: lessons from animal models
Erratum in
- Front Immunol. 2014;5:407. Grassi, Fabio [added]
Abstract
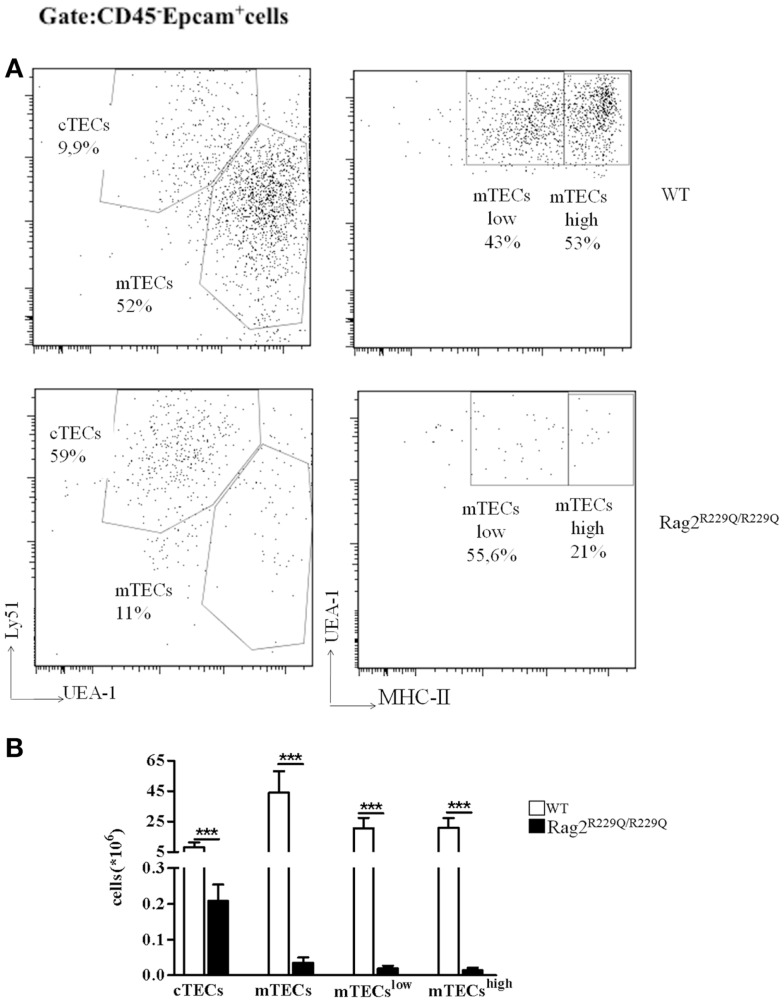
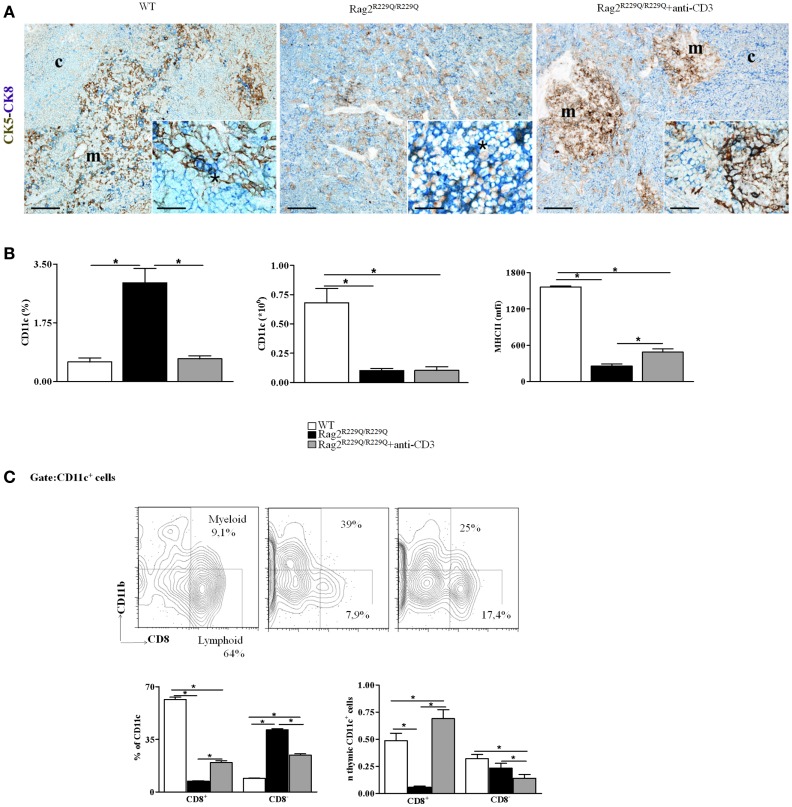
Thymocytes and thymic epithelial cells (TECs) cross-talk is essential to support T cell development and preserve thymic architecture and maturation of TECs and Foxp3(+) natural regulatory T cells. Accordingly, disruption of thymic lymphostromal cross-talk may have major implications on the thymic mechanisms that govern T cell tolerance. Several genetic defects have been described in humans that affect early stages of T cell development [leading to severe combined immune deficiency (SCID)] or late stages in thymocyte maturation (resulting in combined immunodeficiency). Hypomorphic mutations in SCID-causing genes may allow for generation of a limited pool of T lymphocytes with a restricted repertoire. These conditions are often associated with infiltration of peripheral tissues by activated T cells and immune dysregulation, as best exemplified by Omenn syndrome (OS). In this review, we will discuss our recent findings on abnormalities of thymic microenvironment in OS with a special focus of defective maturation of TECs, altered distribution of thymic dendritic cells and impairment of deletional and non-deletional mechanisms of central tolerance. Here, taking advantage of mouse models of OS and atypical SCID, we will discuss how modifications in stromal compartment impact and shape lymphocyte differentiation, and vice versa how inefficient T cell signaling results in defective stromal maturation. These findings are instrumental to understand the extent to which novel therapeutic strategies should act on thymic stroma to achieve full immune reconstitution.
Keywords: Omenn and leaky SCID models; Rag deficiency; central tolerance; thymic cross-talk; thymic reconstitution; thymus.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources