

Brain serotonin determines maternal behavior and offspring survival
- PMID: 25077934
- PMCID: PMC4804711
- DOI: 10.1111/gbb.12159
Brain serotonin determines maternal behavior and offspring survival
Abstract
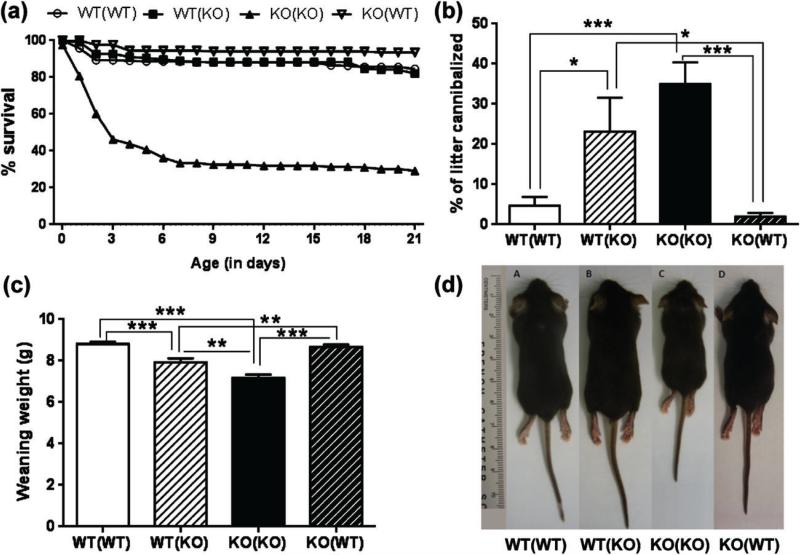
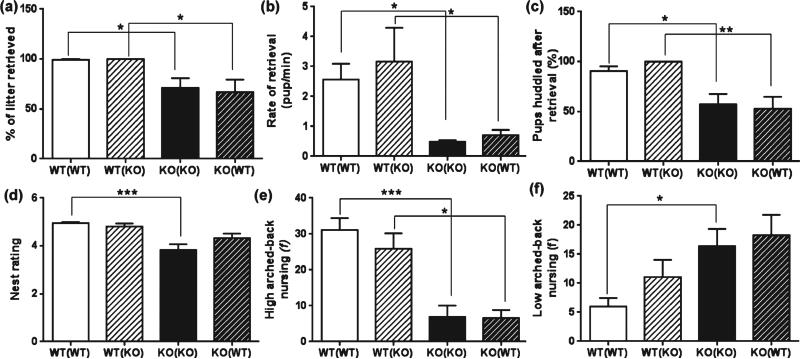
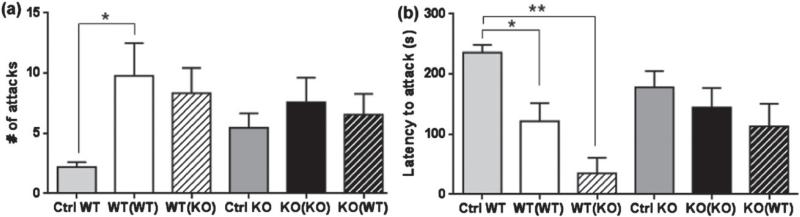
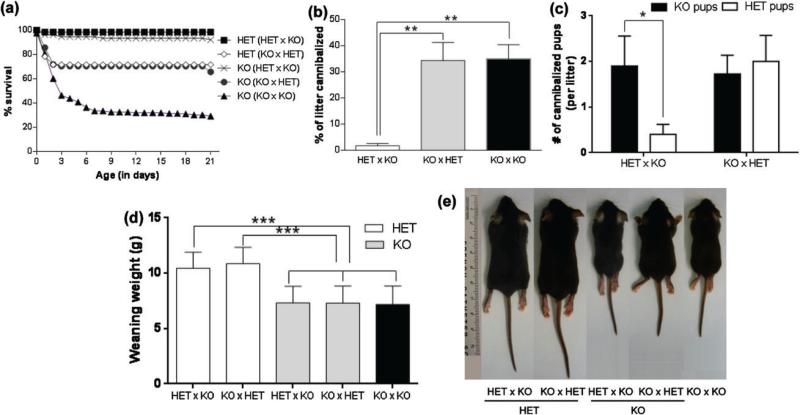
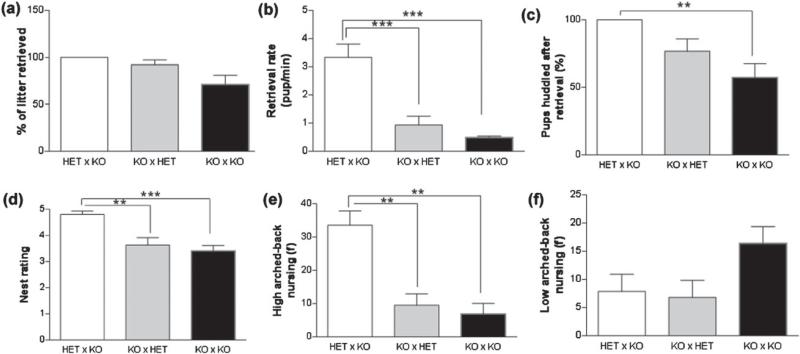
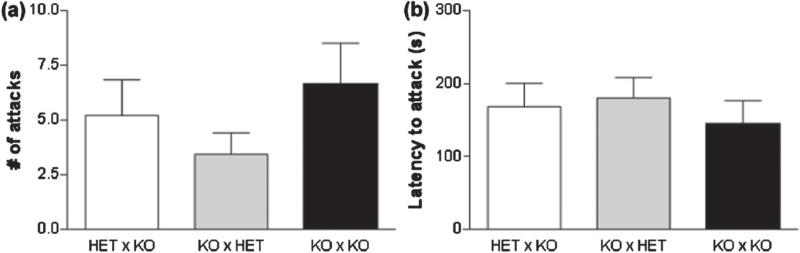
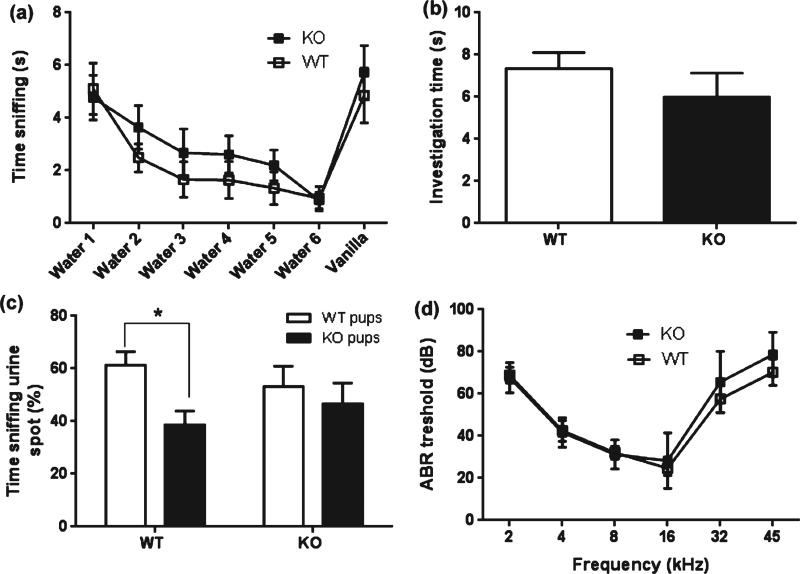
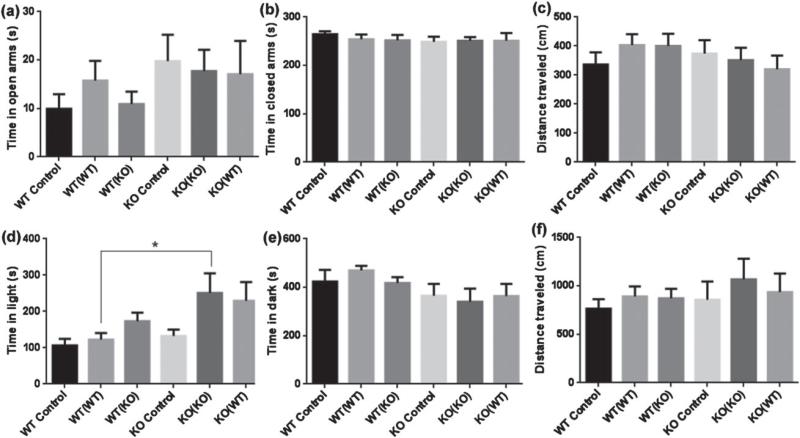
Maternal care is an indispensable component of offspring survival and development in all mammals and necessary for reproductive success. Although brain areas regulating maternal behaviors are innervated by serotonergic afferents, very little is known about the role of this neurotransmitter in these behaviors. To evaluate the contribution of serotonin to maternal care, we used mice with a null mutation in the gene for tryptophan hydroxylase-2 (TPH2), which results in a genetic depletion of brain serotonin, and tested them in a wide range of maternal behavior paradigms. We found that litters born to and reared by TPH2(-/-) mothers showed decreased survival, lower weaning weights and increased cannibalization. In addition, TPH2(-/-) mothers performed poorly in pup retrieval, huddling, nest construction and high-arched back nursing. Aggression in TPH2(-/-) dams was not triggered by lactation and was steadily high. Survival and weaning weight deficits of TPH2(-/-) pups were rescued by cross-fostering and in litters of mixed genotype (TPH2(-/-) and TPH2(-/+) ). However, the maternal behaviors of TPH2(-/-) dams did not improve when rearing either TPH2(+/+) pups or mixed-genotype litters. In addition, TPH2(-/-) pups significantly worsened the behavior of TPH2(+/+) dams with respect to cannibalism, weaning weight and latency to attack. Olfactory and auditory functions of TPH2(-/-) females or anxiety-like behaviors did not account for these maternal alterations as they were equal to their TPH2(+/+) counterparts. These findings illustrate a profound influence of brain serotonin on virtually all elements of maternal behavior and establish that TPH2(-/-) pups can engender maladaptive mothering in dams of both genotypes.
Keywords: Maternal behavior; TPH2; TPH2 knockout; pup survival; serotonin.
© 2014 John Wiley & Sons Ltd and International Behavioural and Neural Genetics Society.
Figures
References
-
- Barofsky AL, Taylor J, Tizabi Y, Kumar R, Jones-Quartey K. Specific neurotoxin lesions of median raphe serotonergic neurons disrupt maternal behavior in the lactating rat. Endocrinology. 1983;113:1884–1893. - PubMed
-
- Bosch OJ. Maternal nurturing is dependent on her innate anxiety: the behavioral roles of brain oxytocin and vasopressin. Horm Behav. 2011;59:202–212. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
