

Phagocytic uptake of oxidized heme polymer is highly cytotoxic to macrophages
- PMID: 25078090
- PMCID: PMC4117526
- DOI: 10.1371/journal.pone.0103706
Phagocytic uptake of oxidized heme polymer is highly cytotoxic to macrophages
Abstract
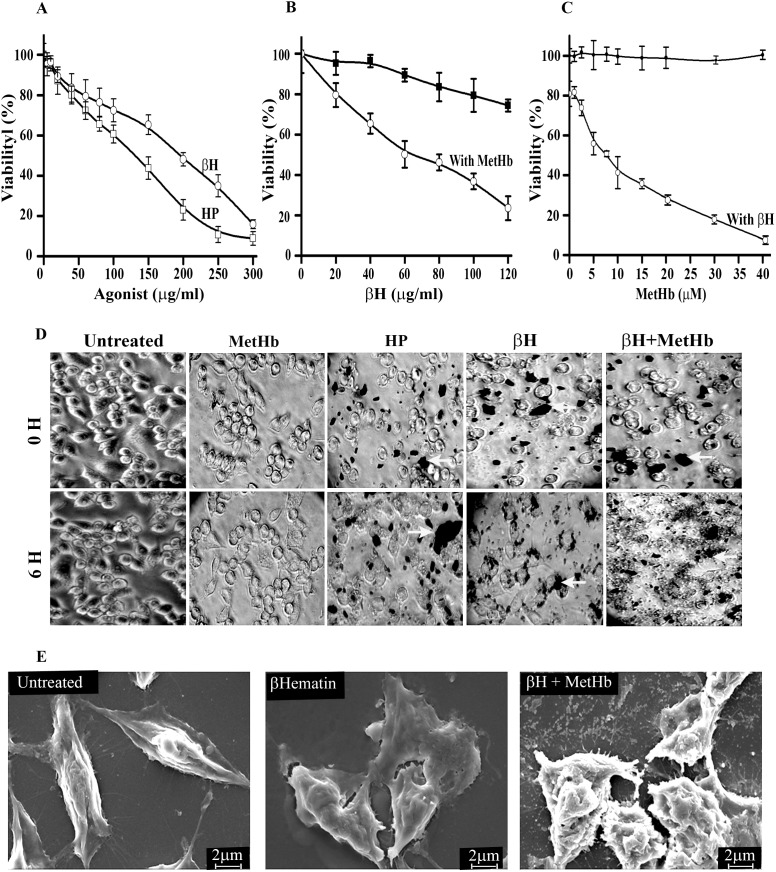
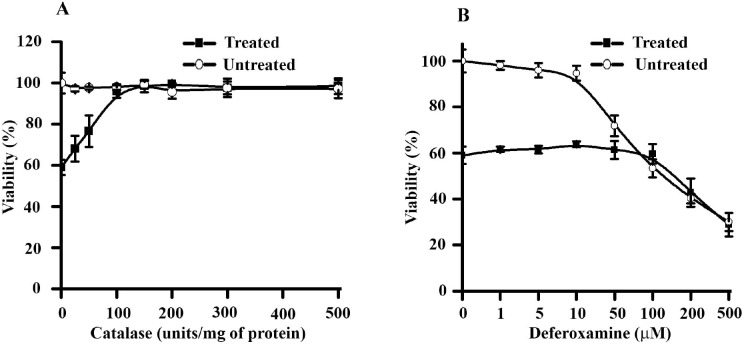
Apoptosis in macrophages is responsible for immune-depression and pathological effects during malaria. Phagocytosis of PRBC causes induction of apoptosis in macrophages through release of cytosolic factors from infected cells. Heme polymer or β-hematin causes dose-dependent death of macrophages with LC50 of 132 µg/ml and 182 µg/ml respectively. The toxicity of hemin or heme polymer was amplified several folds in the presence of non-toxic concentration of methemoglobin. β-hematin uptake in macrophage through phagocytosis is crucial for enhanced toxicological effects in the presence of methemoglobin. Higher accumulation of β-hematin is observed in macrophages treated with β-hematin along with methemoglobin. Light and scanning electron microscopic observations further confirm accumulation of β-hematin with cellular toxicity. Toxicological potentiation of pro-oxidant molecules toward macrophages depends on generation of H2O2 and independent to release of free iron from pro-oxidant molecules. Methemoglobin oxidizes β-hematin to form oxidized β-hematin (βH*) through single electron transfer mechanism. Pre-treatment of reaction mixture with spin-trap Phenyl-N-t-butyl-nitrone dose-dependently reverses the β-hematin toxicity, indicates crucial role of βH* generation with the toxicological potentiation. Acridine orange/ethidium bromide staining and DNA fragmentation analysis indicate that macrophage follows an oxidative stress dependent apoptotic pathway to cause death. In summary, current work highlights mutual co-operation between methemoglobin and different pro-oxidant molecules to enhance toxicity towards macrophages. Hence, methemoglobin peroxidase activity can be probed for subduing cellular toxicity of pro-oxidant molecules and it may in-turn make up for host immune response against the malaria parasite.
Conflict of interest statement
Figures




References
-
- Sachs J, Malaney P (2002) The economic and social burden of malaria. Nature 415: 680–685. - PubMed
-
- Pamplona A, Hanscheid T, Epiphanio S, Mota MM, Vigario AM (2009) Cerebral malaria and the hemolysis/methemoglobin/heme hypothesis: shedding new light on an old disease. Int J Biochem Cell Biol 41: 711–716. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
