

Multiplicity of 5' cap structures present on short RNAs
- PMID: 25079783
- PMCID: PMC4117478
- DOI: 10.1371/journal.pone.0102895
Multiplicity of 5' cap structures present on short RNAs
Abstract
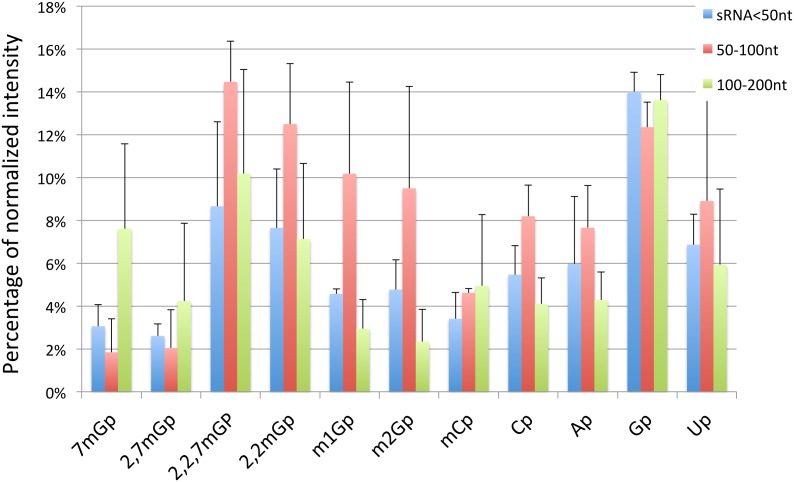
Most RNA molecules are co- or post-transcriptionally modified to alter their chemical and functional properties to assist in their ultimate biological function. Among these modifications, the addition of 5' cap structure has been found to regulate turnover and localization. Here we report a study of the cap structure of human short (<200 nt) RNAs (sRNAs), using sequencing of cDNA libraries prepared by enzymatic pretreatment of the sRNAs with cap sensitive-specificity, thin layer chromatographic (TLC) analyses of isolated cap structures and mass spectrometric analyses for validation of TLC analyses. Processed versions of snoRNAs and tRNAs sequences of less than 50 nt were observed in capped sRNA libraries, indicating additional processing and recapping of these annotated sRNAs biotypes. We report for the first time 2,7 dimethylguanosine in human sRNAs cap structures and surprisingly we find multiple type 0 cap structures (mGpppC, 7mGpppG, GpppG, GpppA, and 7mGpppA) in RNA length fractions shorter than 50 nt. Finally, we find the presence of additional uncharacterized cap structures that wait determination by the creation of needed reference compounds to be used in TLC analyses. These studies suggest the existence of novel biochemical pathways leading to the processing of primary and sRNAs and the modifications of their RNA 5' ends with a spectrum of chemical modifications.
Conflict of interest statement
Figures
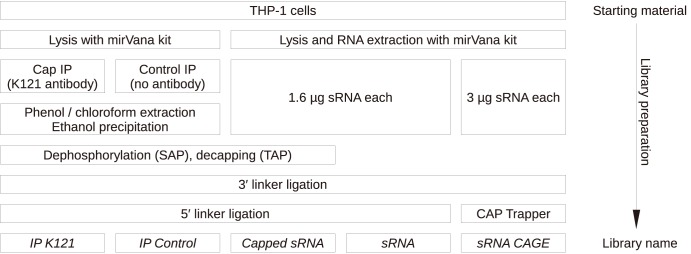
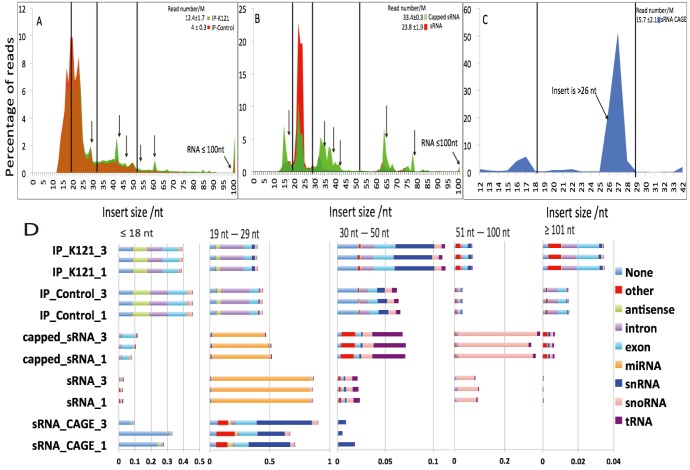
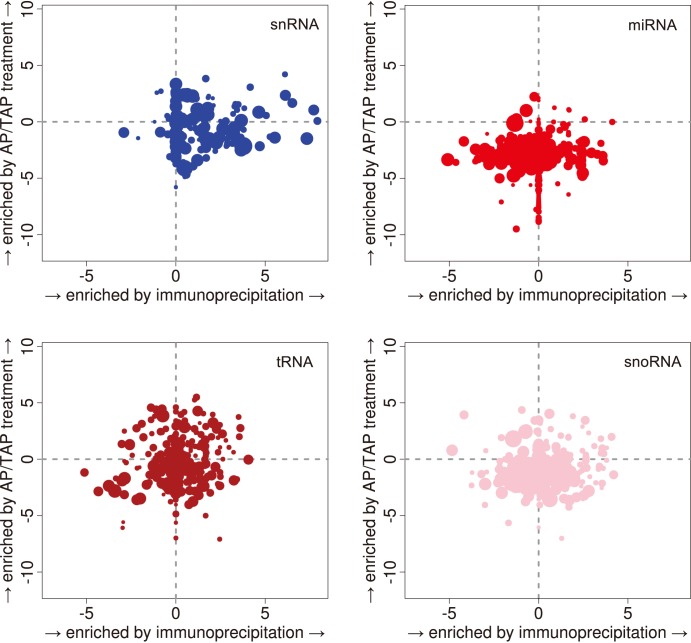
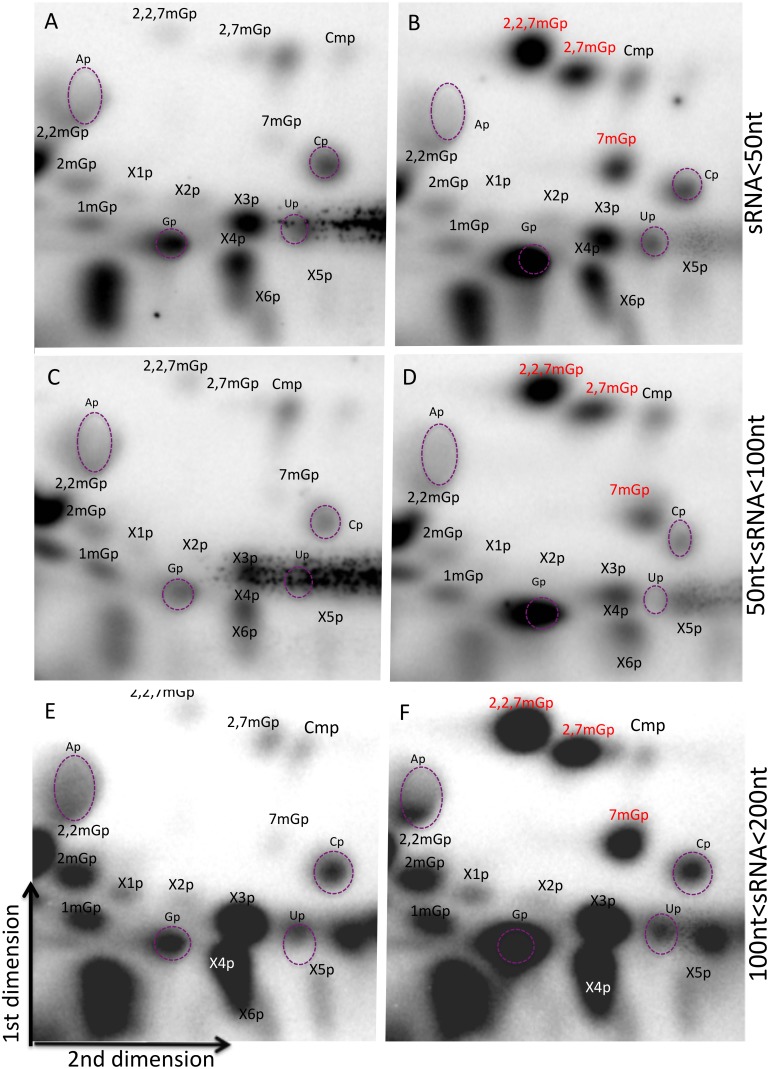
References
-
- Matera AG, Terns RM, Terns MP (2007) Non-coding RNAs: lessons from the small nuclear and small nucleolar RNAs. Nat Rev Mol Cell Biol 8: 209–220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
