

Seasonal oscillation of liver-derived hibernation protein complex in the central nervous system of non-hibernating mammals
- PMID: 25079892
- PMCID: PMC4117459
- DOI: 10.1242/jeb.095976
Seasonal oscillation of liver-derived hibernation protein complex in the central nervous system of non-hibernating mammals
Abstract
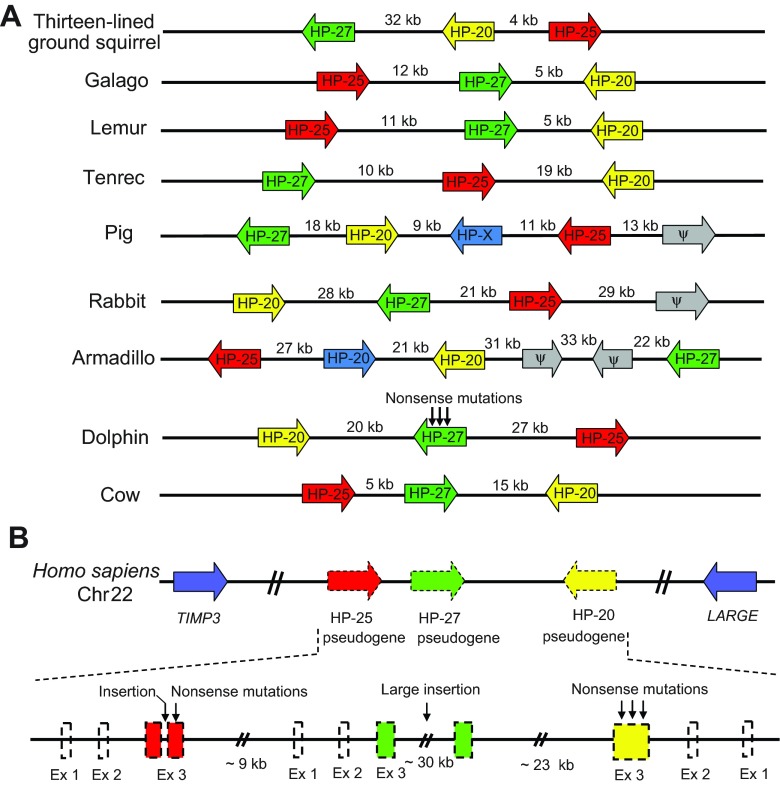
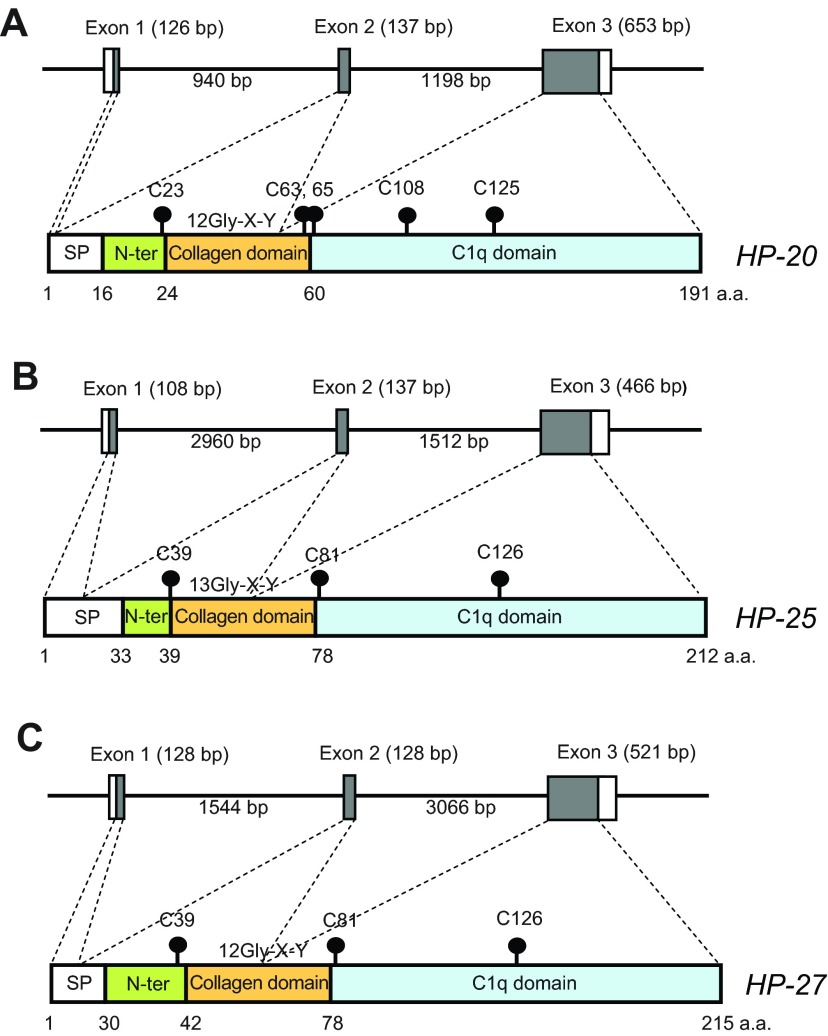
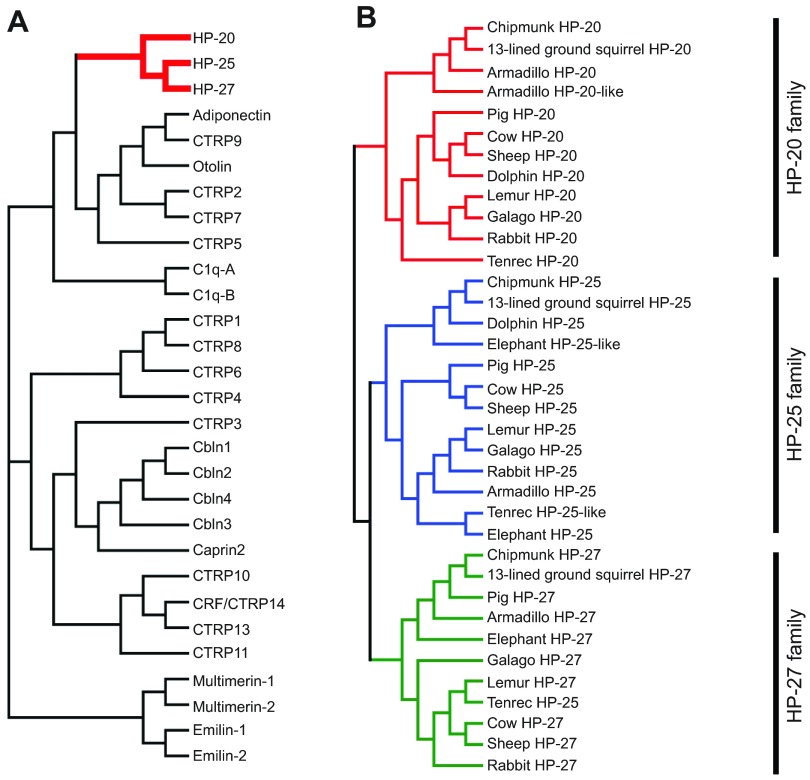
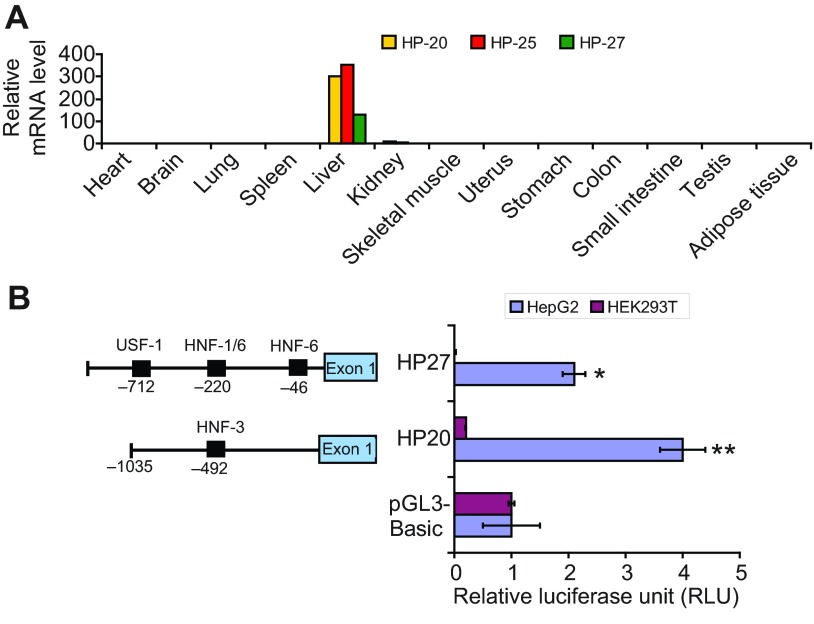
Mammalian hibernation elicits profound changes in whole-body physiology. The liver-derived hibernation protein (HP) complex, consisting of HP-20, HP-25 and HP-27, was shown to oscillate circannually, and this oscillation in the central nervous system (CNS) was suggested to play a role in hibernation. The HP complex has been found in hibernating chipmunks but not in related non-hibernating tree squirrels, leading to the suggestion that hibernation-specific genes may underlie the origin of hibernation. Here, we show that non-hibernating mammals express and regulate the conserved homologous HP complex in a seasonal manner, independent of hibernation. Comparative analyses of cow and chipmunk HPs revealed extensive biochemical and structural conservations. These include liver-specific expression, assembly of distinct heteromeric complexes that circulate in the blood and cerebrospinal fluid, and the striking seasonal oscillation of the HP levels in the blood and CNS. Central administration of recombinant HPs affected food intake in mice, without altering body temperature, physical activity levels or energy expenditure. Our results demonstrate that HP complex is not unique to the hibernators and suggest that the HP-regulated liver-brain circuit may couple seasonal changes in the environment to alterations in physiology.
Keywords: CTRP; Cerebrospinal fluids; Hibernation; Metabolism; Plasma proteins.
© 2014. Published by The Company of Biologists Ltd.
Figures

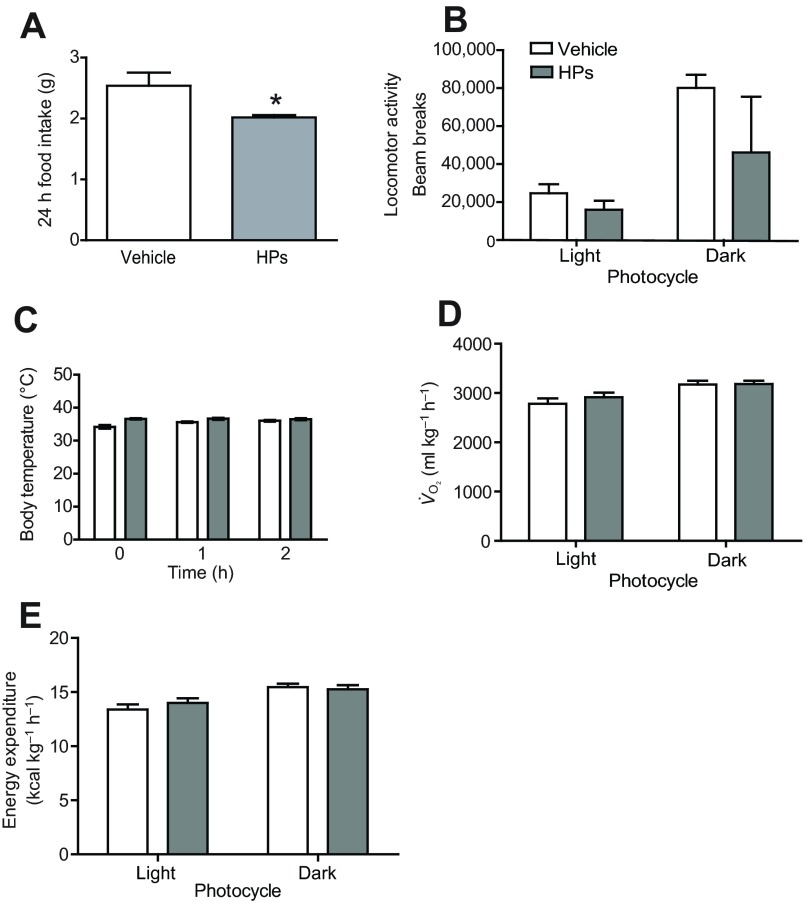
 (D) and energy expenditure (E). *P<0.05 (between vehicle and recombinant HPs).
(D) and energy expenditure (E). *P<0.05 (between vehicle and recombinant HPs).References
-
- Andrews M. T. (2007). Advances in molecular biology of hibernation in mammals. Bioessays 29, 431-440 - PubMed
-
- Ben Saad M., Baylé J. D. (1985). Seasonal changes in plasma testosterone, thyroxine, and cortisol levels in wild rabbits (Oryctolagus cuniculus algirus) of Zembra island. Gen. Comp. Endocrinol. 57, 383-388 - PubMed
-
- Berg A. H., Combs T. P., Du X., Brownlee M., Scherer P. E. (2001). The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 7, 947-953 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
