

Review
doi: 10.1007/s00018-014-1695-z.
Epub 2014 Aug 1.
Common and unique features of viral RNA-dependent polymerases
Affiliations
- PMID: 25080879
- PMCID: PMC4207942
- DOI: 10.1007/s00018-014-1695-z
Item in Clipboard
Review
Common and unique features of viral RNA-dependent polymerases
Cell Mol Life Sci.
2014 Nov.
Abstract
Eukaryotes and bacteria can be infected with a wide variety of RNA viruses. On average, these pathogens share little sequence similarity and use different replication and transcription strategies. Nevertheless, the members of nearly all RNA virus families depend on the activity of a virally encoded RNA-dependent polymerase for the condensation of nucleotide triphosphates. This review provides an overview of our current understanding of the viral RNA-dependent polymerase structure and the biochemistry and biophysics that is involved in replicating and transcribing the genetic material of RNA viruses.
Figures
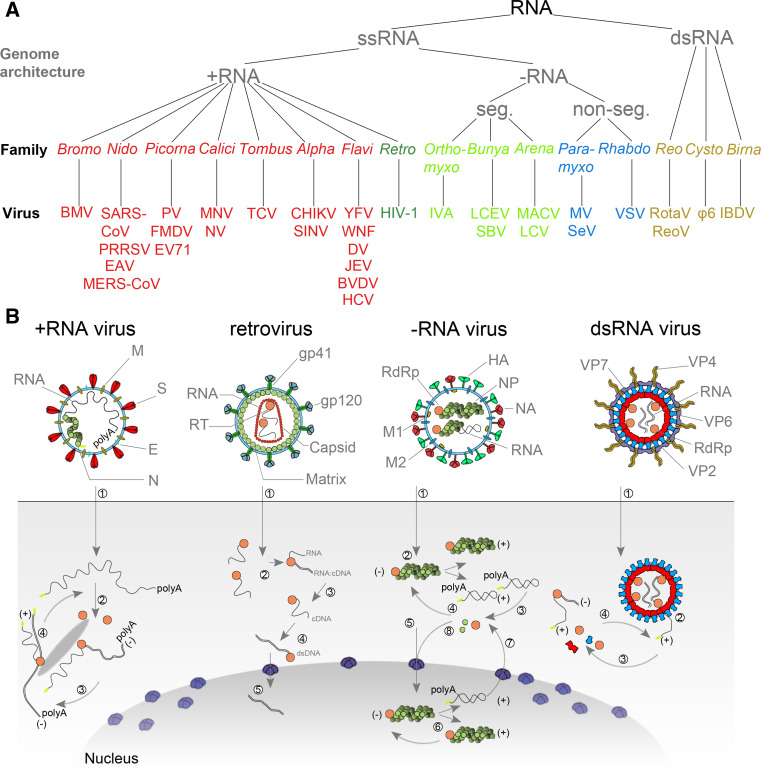
Taxonomy and replication strategies of RNA viruses. a Simplified taxonomy of the genome architecture of the RNA viruses described in this review. See main text for used abbreviations. b (+RNA virus) Infection with a +RNA virus—as exemplified here with a CoV-like virion—releases a single-stranded RNA genome into the cytoplasm (1) [81, 173, 174]. (2) Translation of the 5′-terminal open-reading frame of the genome produces the viral replicase. (3) This multi-enzyme complex includes RdRp activity (orange) and associates with intracellular membranes before −RNA synthesis commences. Newly synthesised −RNAs are subsequently used to produce new +RNAs (4), which are typically capped (yellow) and polyadenylated (polyA). (Retrovirus) HIV-1 genomes are packaged as ssRNA in virions. When the ssRNA is released (1) a cDNA copy is synthesised by the RT (2). The RNA is next degraded by the intrinsic RNase H activity in the RT (3) and the single stranded cDNA converted to dsDNA (4). The dsDNA is imported in the nucleus (5) for integration into the host’s genetic material. (−RNA virus) (1) As illustrated here with an IAV-like particle, infection with an −RNA virus releases a viral RNA genome that is associated with a viral polymerase (orange) and nucleoprotein (green). (2) In the case of non-segmented −RNA viruses, these complexes support transcription to produce viral mRNAs or cRNAs. (3) Viral mRNAs are next translated and new viral proteins complex with cRNAs to synthesise new vRNAs. (5) The vRNA-containing complexes of some segmented −RNA viruses are imported into the nucleus of the host cell, where (6) the RdRp produces mRNAs or cRNAs. (7) mRNAs are transported to the cytoplasm, while cRNAs are bound by new viral proteins to form cRNPs for −RNA synthesis. (dsRNA virus) Fully duplexed RNA genomes lack cap and polyA elements. (1) The RdRp (orange), therefore, transcribes the viral genome inside the capsid of the virion (blue and red), so viral mRNAs can be (2) released into the cytoplasm as illustrated here with a rotavirus-like virion. In the cytoplasm the mRNA is translated (3) or replicated by newly synthesised viral RdRps (4) [175, 176]
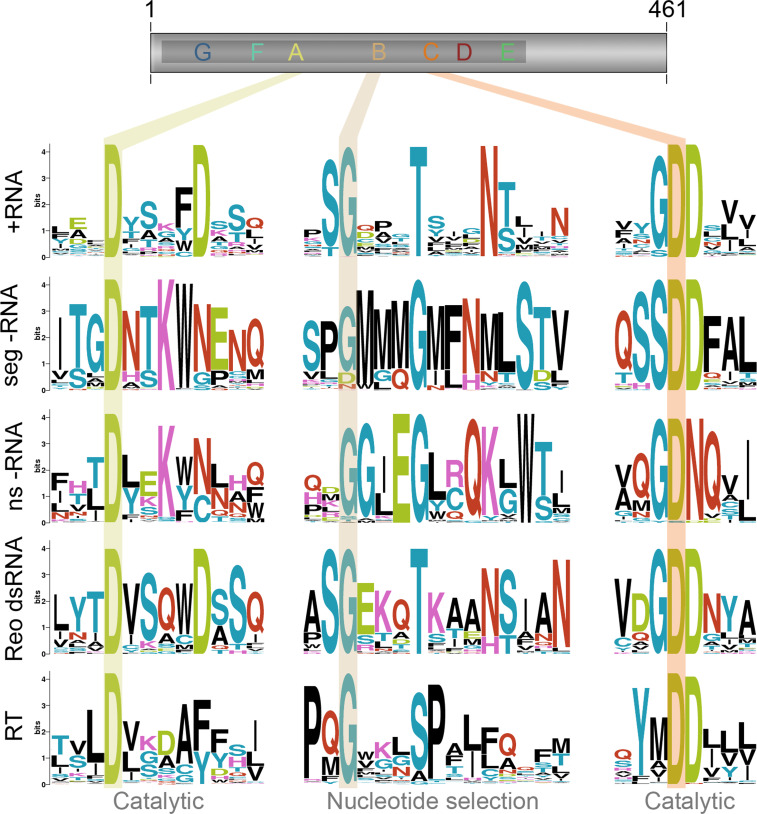
Key conserved residues of the RNA polymerase domain. Motifs A–C reside in the middle of the typical RNA-dependent polymerase domain as shown here in the schematic of the poliovirus 3Dpol subunit. They are involved in catalysis and nucleotide selection and the residues involved in these processes are highly conserved. The key residues of these motifs are shaded across the RNA polymerase domains of positive strand RNA viruses (+RNA), segmented negative strand RNA viruses (seg −RNA), non-segmented negative strand RNA viruses (ns −RNA), double strand RNA viruses of the reovirus family (Reo dsRNA), and reverse transcriptases (RT). Sequence logo images were created using prosite accession numbers PDSC50507 and PDOC50878
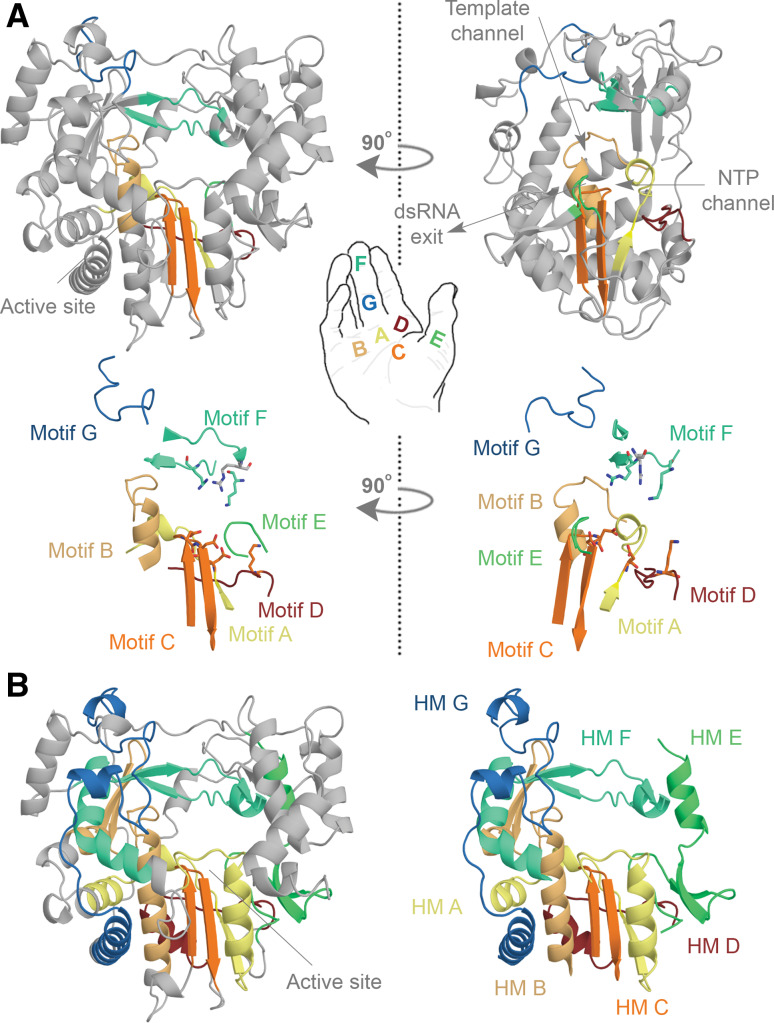
Conserved structural elements in the RNA virus polymerase. a Structure of the FMDV RdRp. The motifs A, B, C, D, E, F, and G are colour coded yellow, gold, orange, red, light green, aquamarine, and blue. Overall the polymerase structure adopts a shape that resembles a cupped right hand. Herein, motifs A–E lie on the palm, while motif F and G are part of the fingers. In the side-view of the enzyme the location of the template and NTP channels is indicated. b Conserved structural elements of the FMDV RdRp. Homomorphs A–G were mapped according to Ref. [15] and colour coded yellow, gold, orange, red, light green, aquamarine, and blue, respectively. Images A and B are based on PDB accession 2E9R
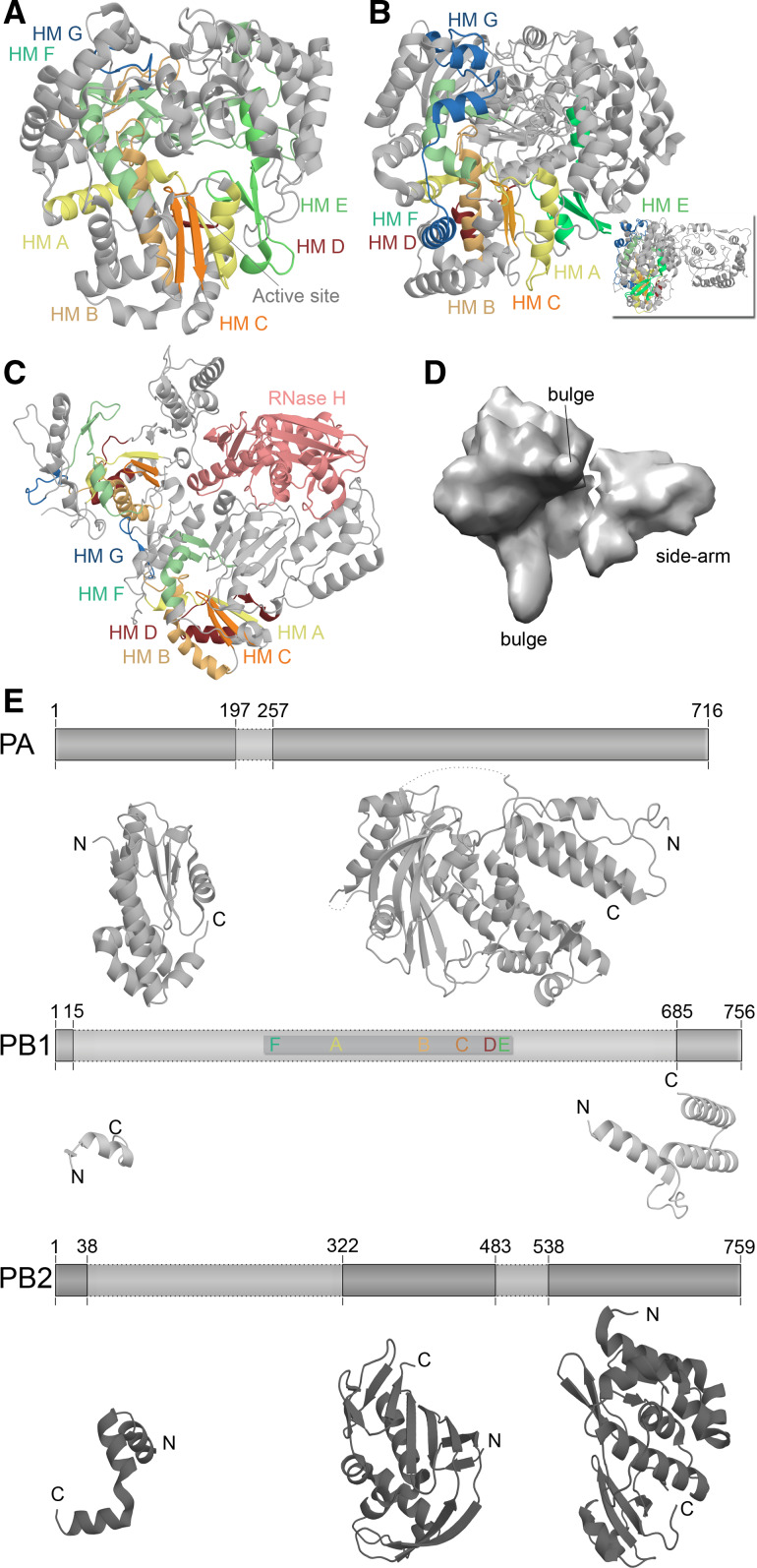
Structural differences among RNA virus polymerases. a Structure of the ϕ6 RdRp P2 based on PDB accession 1HI0. b Structure of the JEV polymerase based on PBD entry 4K6 M. Inset depicts 90° rotation of polymerase to visualise the N-terminal methyltransferase domain. c Structure of the HIV-1 RT based on PDB accession 3V4I. The RT is comprised of the p66 (left) and p51 (right) protein subunits. Only the p66 subunit has an RNase domain (pink). Homomorphs A–G are colour coded yellow, gold, orange, red, light green, aquamarine, and blue, respectively in Fig. 4a–c. d EM model of the IAV RdRp based on PDBe entry EMD-2213. Structural features were identified by Moeller et al. [79]. e The IAV polymerase consists of the subunits PA, PB1, and PB2. Six of the seven canonical RNA-dependent polymerase domains motifs are found in PB1, which are colour coded as in Fig. 3. Presently only significant structural information is available for PA and PB2. Figure based on PBD entries 2VY6, 2W69, 2ZNL, 2ZTT, 3EBJ, and 4CB4
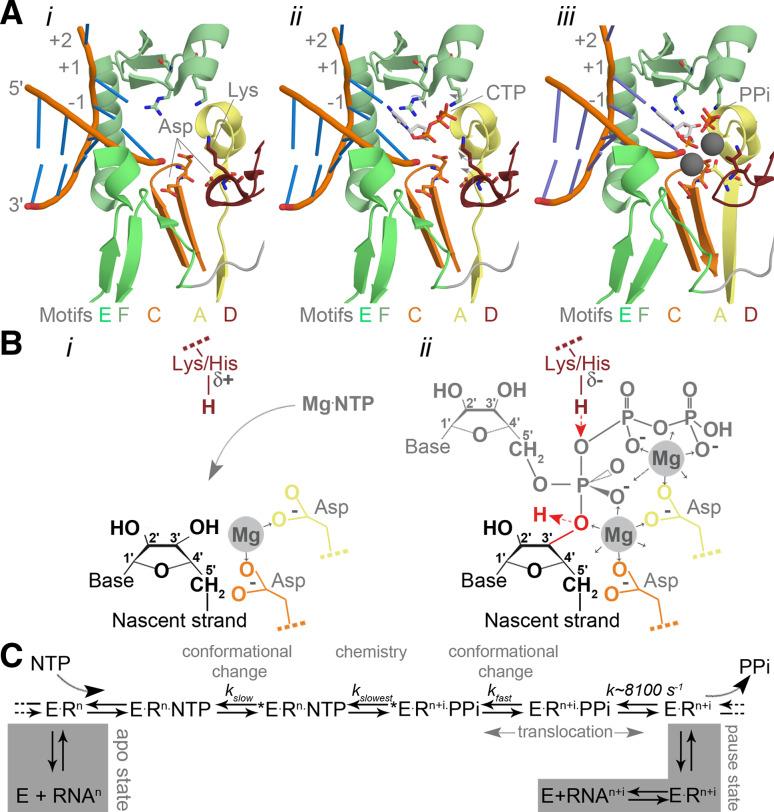
Catalysis in the RNA virus polymerase active site. a Structure of the PV active site as it moves from a native state or elongation complex (i) to an open complex (ii), and a closed complex (iii). The closed complex depicted here shows the active site after catalysis. Highlighted are the metal-binding aspartates of motifs A and C, and the lysine of motif D that acts as general acid. Colour coding by motif as in Fig. 3. Image based on PDB accessions 3Ol6, 3OLB, and 3OL7. b Schematic presentation of the RdRp active site. The aspartates (Asp) of motif A (yellow) and C (orange) bind divalent metal ions (marked Mg and shaded grey), which are used to coordinate the formation of a new phosphodiester bond at the 3′-OH (red in panel ii) of the nascent strand (yellow). The general acid (red Lys/His in panel ii) is positioned near the β-phosphate of the incoming NTP to protonate the PPi leaving group. c Simplified schematic of the kinetic steps of RNA polymerases. Asterisk indicates closed complex
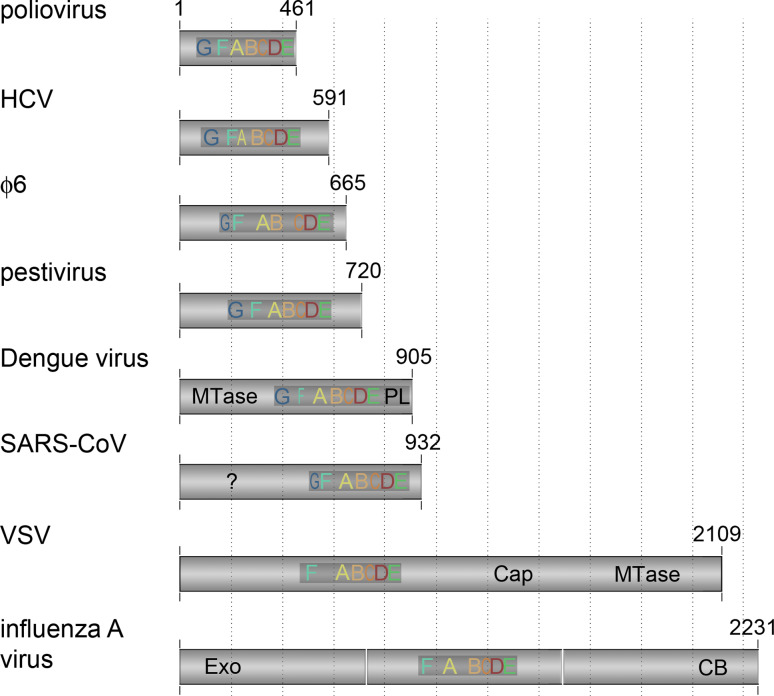
Sizes and additional domains of RNA virus polymerases. The RNA-dependent polymerase domain consists of approximately 400 amino acids. The size of the rest of the subunit varies significantly among RNA-dependent polymerases, as does the conservation of motifs F and G. The additional sequences of the polymerase subunit often contain functional domains, such as a methyltransferase (MTase), exonuclease (Exo), capping domain (Cap), or cap-binding domain (CB). The SAR-CoV RdRp has an additional N-terminal domain that is conserved, but to which no function has been ascribed yet (?). Figure based on alignments from references [15, 39, 83, 177, 178]
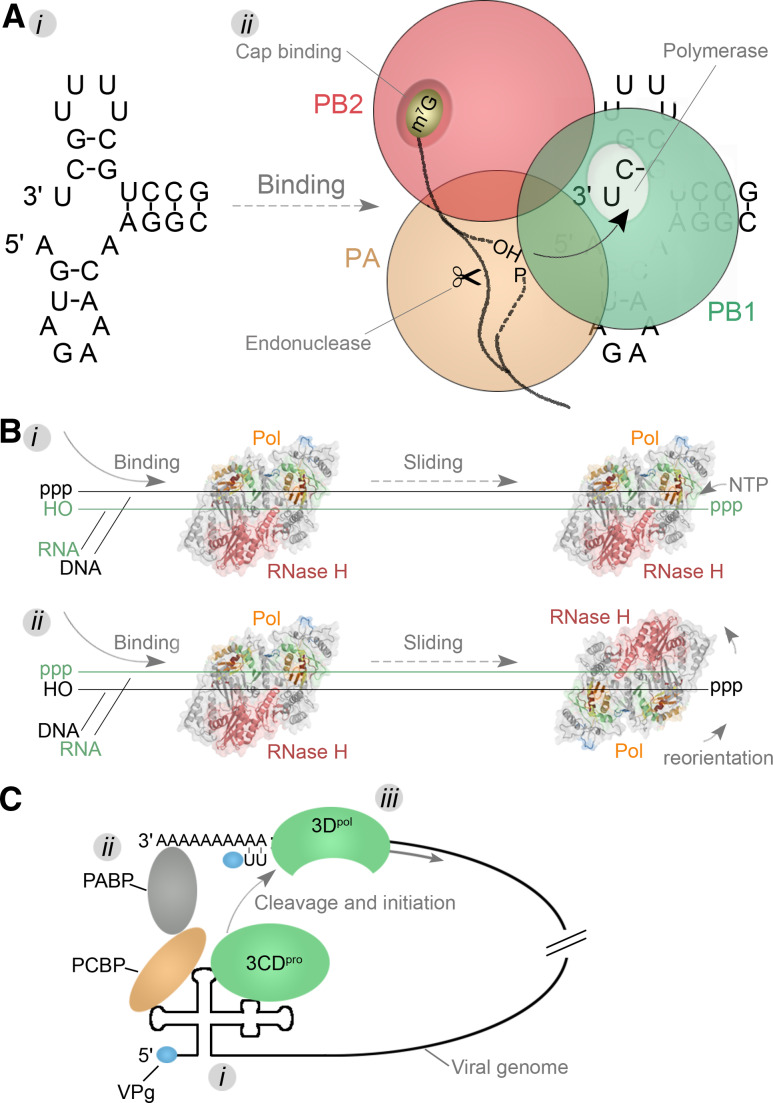
Template recognition mechanisms. a
i The IAV RNA promoter forms through hybridization of the terminal ends of a viral genome segment. ii The structure is next bound by the heterotrimeric RdRp complex that consists of subunits PB1 (green), PB2 (red), and PA (orange). During transcription, the PB2 subunit binds the cap of pre-mRNAs. These pre-mRNAs are next cleaved by the PA subunit and transferred to the PB1 subunit. Here nucleotide condensation takes place on the 3′-OH of the capped primers. b The HIV-1 RT is comprised of the subunits p66 and p51, of which only the p66 subunit has an active polymerase (colour coded as Fig. 4) and RNase domain (pink). The enzyme can bind to the template in a random orientation and switch between the activities by tumbling around its axis once a polymerase (i) or RNase (ii) substrate has been encountered. c A model for the circularisation of the PV genome. First viral proteins and the 3CDpro cleavage intermediate bind to conserved 5′ structures on the genome (i). Next a protein bridge is formed that brings together the 5′ and 3′-ends of the genome (ii). This step subsequently ensures that the viral protease is activated and the viral polymerase 3Dpol is released in close proximity to the 3′-end of the viral genome. Here it can initiate -RNA synthesis using VPg as protein primer
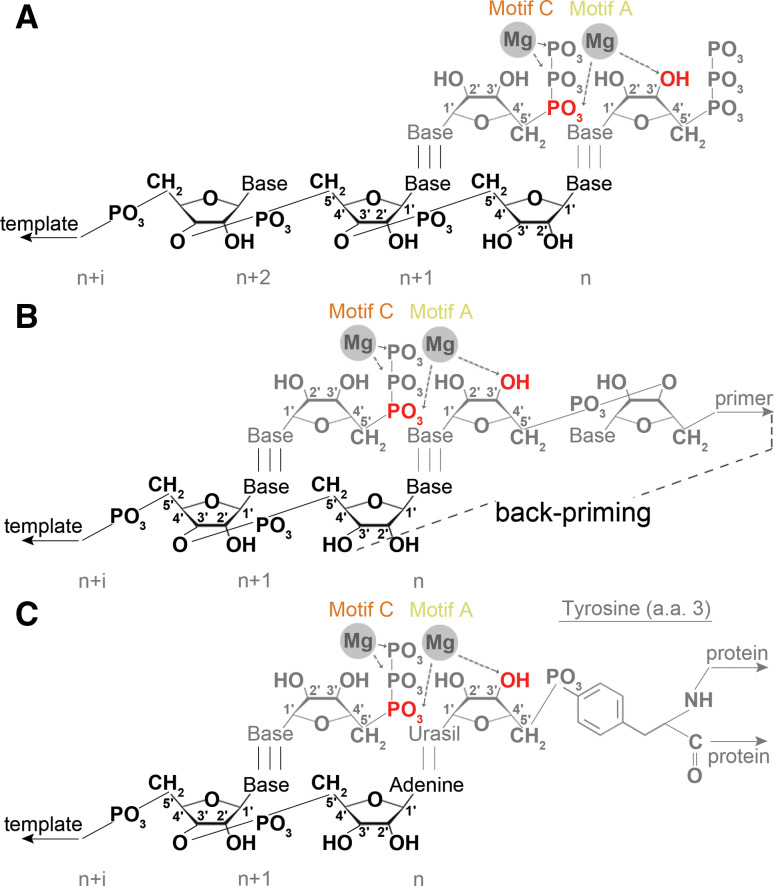
Modes of initiation. schematic presentation of a de novo initiation, b primer-dependent initiation, and c protein-dependent initiation. In the latter scenario, the tyrosine residue that is present at position 3 of VPg is used as primer
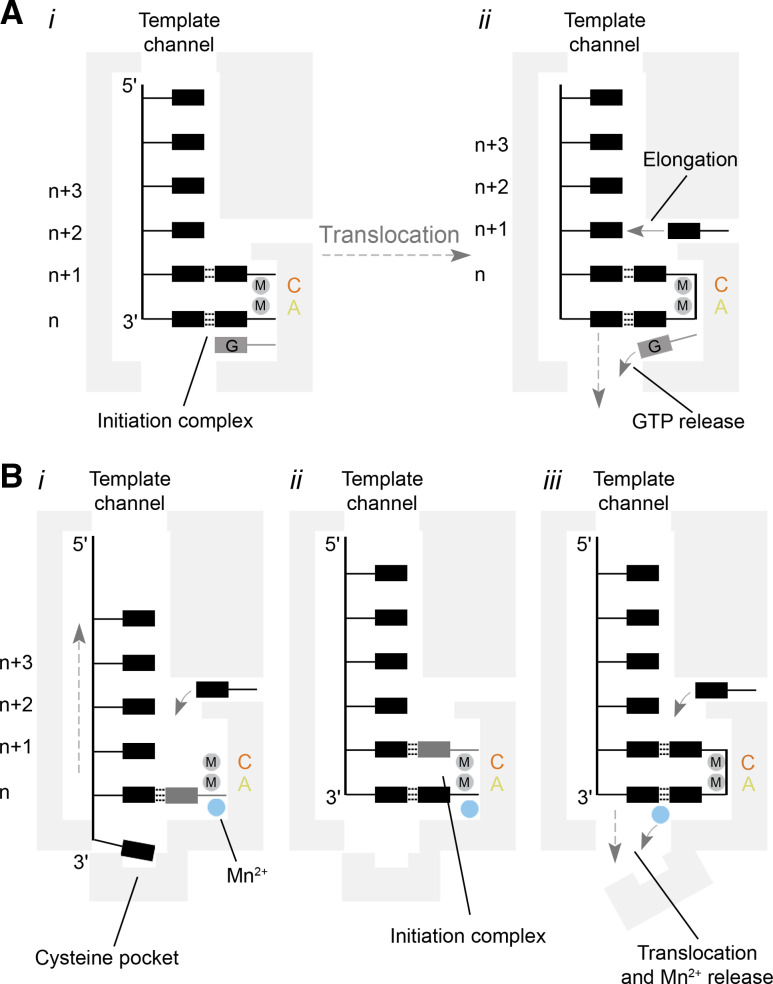
Models for template recognition and de novo initiation by the BVDV and ϕ6 polymerases. a Model of de novo RNA synthesis by a BVDV RdRp. The RdRp binds to the 3′-end of the viral genome (i). Then the template, initiating NTPs, and an additional GTP form an initiation complex that is stabilised by the closed C-terminal loop. After the first polymerase reaction, the GTP is released by opening of the C-terminal loop and translocation of the template–nascent strand duplex (ii). A new NTP can enter the active site via the NTP channel to start elongation. b Simplified model of de novo RNA synthesis by the ϕ6 RdRp. Template recognition occurs through the binding of the 3′-cytosine of the genome to a pyrimidine specificity pocket (i). At the same time, a nucleotide base-pairs with the second base of the template. Its triphosphate group is stabilised by a manganese ion that is coordinated by the palm domain. The 3′-cytosine next translocates to the n-position to base-pair with a second NTP (ii). After nucleotide condensation, the manganese ion triggers a rearrangement of the C-terminal and is subsequently released (iii). In contrast to the GTP in the NS5 RdRp, the manganese ion must be rebound for subsequent elongation
References
-
- Butel JS. Viral carcinogenesis: revelation of molecular mechanisms and etiology of human disease. Carcinogenesis. 2000;21:405–426. - PubMed
-
- Fraaij PL, Bodewes R, Osterhaus AD, Rimmelzwaan GF. The ins and outs of universal childhood influenza vaccination. Future Microbiol. 2011;6:1171–1184. - PubMed
-
- Patel MM, Glass R, Desai R, Tate JE, Parashar UD. Fulfilling the promise of rotavirus vaccines: how far have we come since licensure? Lancet Infect Dis. 2012;12:561–570. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
