

MicroRNA directly enhances mitochondrial translation during muscle differentiation
- PMID: 25083871
- PMCID: PMC4119298
- DOI: 10.1016/j.cell.2014.05.047
MicroRNA directly enhances mitochondrial translation during muscle differentiation
Abstract
MicroRNAs are well known to mediate translational repression and mRNA degradation in the cytoplasm. Various microRNAs have also been detected in membrane-compartmentalized organelles, but the functional significance has remained elusive. Here, we report that miR-1, a microRNA specifically induced during myogenesis, efficiently enters the mitochondria where it unexpectedly stimulates, rather than represses, the translation of specific mitochondrial genome-encoded transcripts. We show that this positive effect requires specific miR:mRNA base-pairing and Ago2, but not its functional partner GW182, which is excluded from the mitochondria. We provide evidence for the direct action of Ago2 in mitochondrial translation by crosslinking immunoprecipitation coupled with deep sequencing (CLIP-seq), functional rescue with mitochondria-targeted Ago2, and selective inhibition of the microRNA machinery in the cytoplasm. These findings unveil a positive function of microRNA in mitochondrial translation and suggest a highly coordinated myogenic program via miR-1-mediated translational stimulation in the mitochondria and repression in the cytoplasm.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
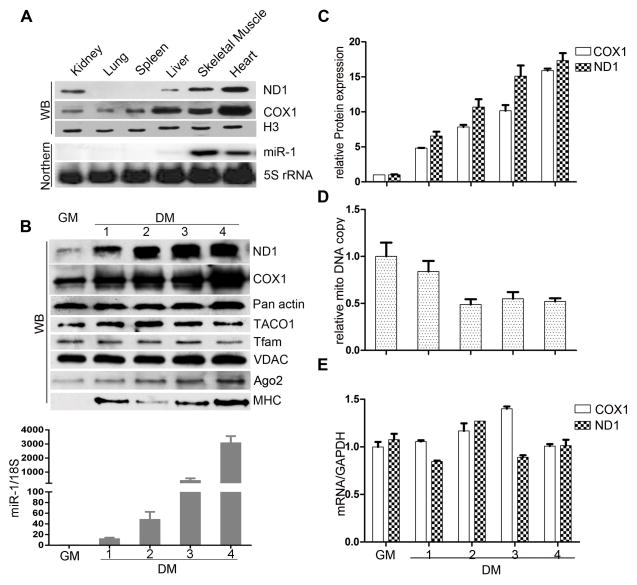
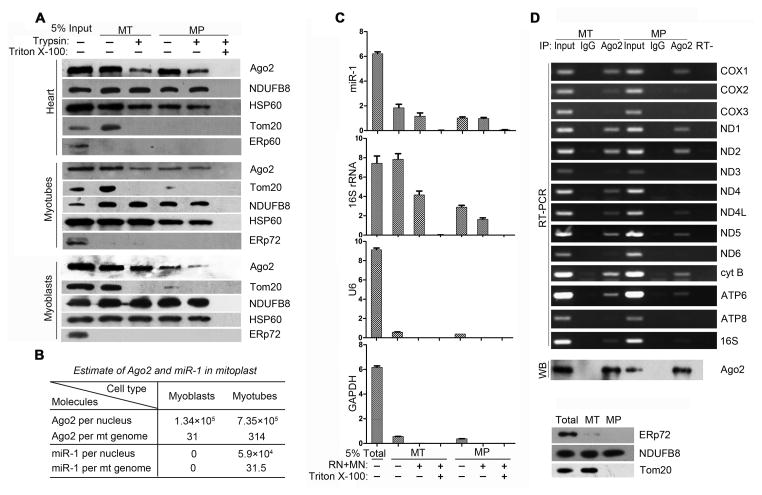
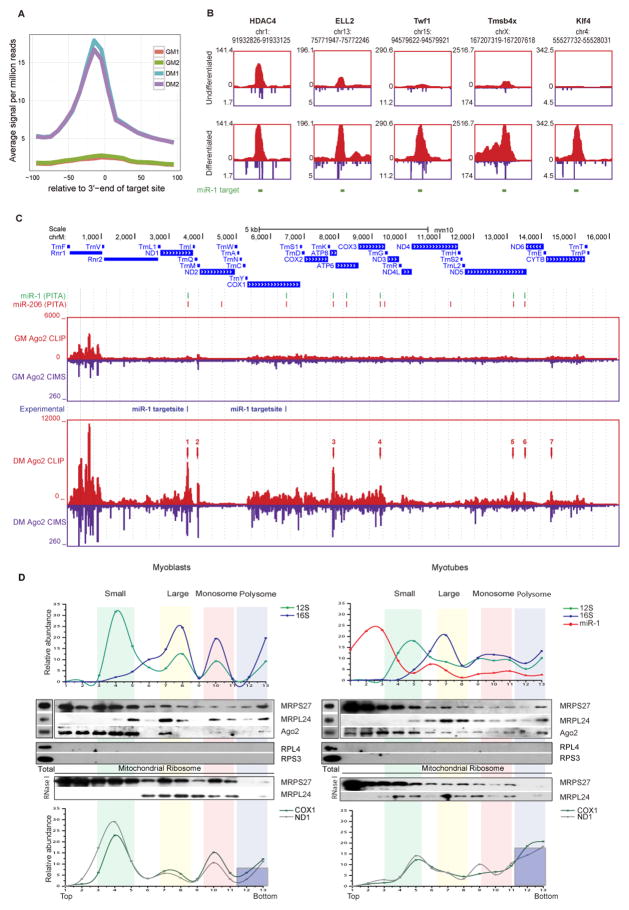
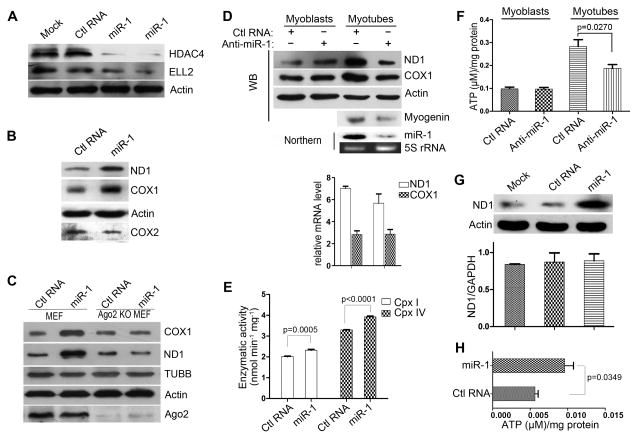
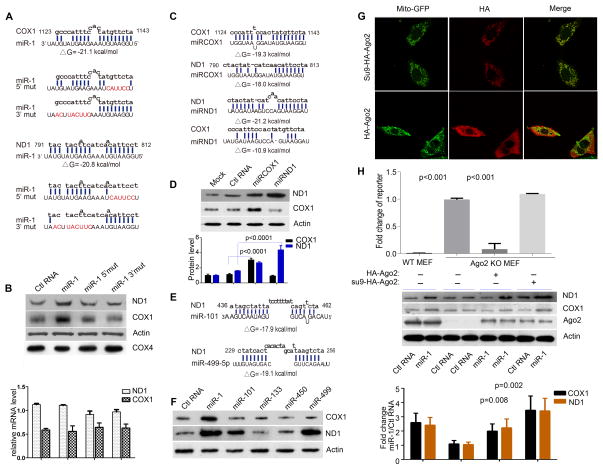
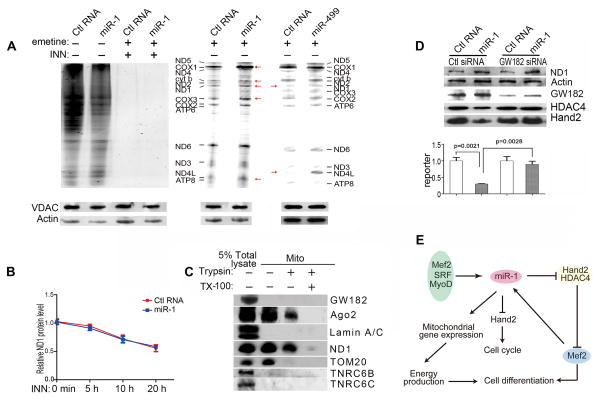
Comment in
-
microManaging mitochondrial translation.Cell. 2014 Jul 31;158(3):477-8. doi: 10.1016/j.cell.2014.07.010. Cell. 2014. PMID: 25083863
References
-
- Antonicka H, Sasarman F, Nishimura T, Paupe V, Shoubridge EA. The mitochondrial RNA-binding protein GRSF1 localizes to RNA granules and is required for posttranscriptional mitochondrial gene expression. Cell metabolism. 2013;17:386–398. - PubMed
-
- Arnold MA, Kim Y, Czubryt MP, Phan D, McAnally J, Qi X, Shelton JM, Richardson JA, Bassel-Duby R, Olson EN. MEF2C transcription factor controls chondrocyte hypertrophy and bone development. Developmental cell. 2007;12:377–389. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
