

Transcription and recombination: when RNA meets DNA
- PMID: 25085910
- PMCID: PMC4107990
- DOI: 10.1101/cshperspect.a016543
Transcription and recombination: when RNA meets DNA
Abstract
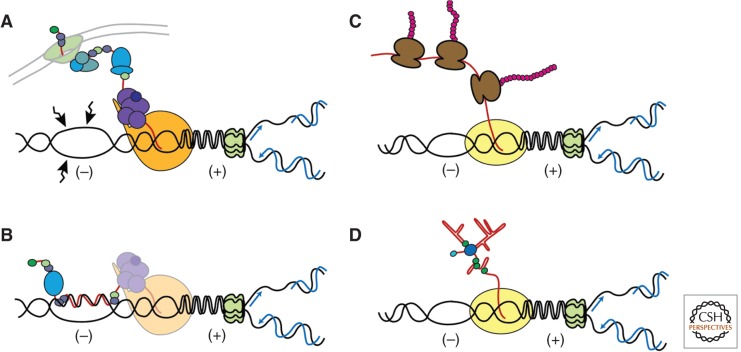
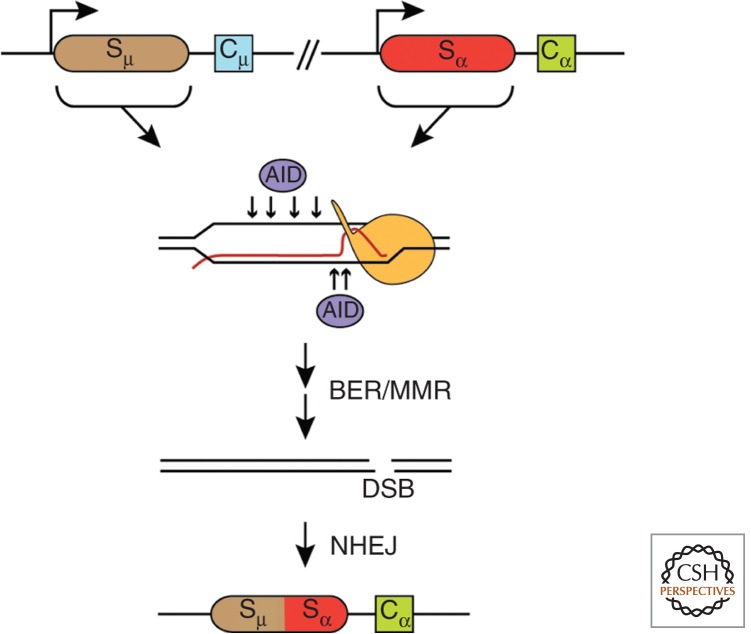
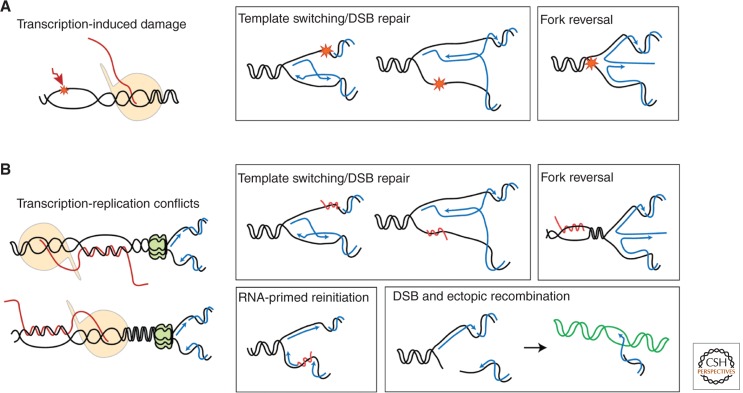
A particularly relevant phenomenon in cell physiology and proliferation is the fact that spontaneous mitotic recombination is strongly enhanced by transcription. The most accepted view is that transcription increases the occurrence of double-strand breaks and/or single-stranded DNA gaps that are repaired by recombination. Most breaks would arise as a consequence of the impact that transcription has on replication fork progression, provoking its stalling and/or breakage. Here, we discuss the mechanisms responsible for the cross talk between transcription and recombination, with emphasis on (1) the transcription-replication conflicts as the main source of recombinogenic DNA breaks, and (2) the formation of cotranscriptional R-loops as a major cause of such breaks. The new emerging questions and perspectives are discussed on the basis of the interference between transcription and replication, as well as the way RNA influences genome dynamics.
Copyright © 2014 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Aguilera A, Garcia-Muse T 2012. R loops: From transcription byproducts to threats to genome stability. Mol Cell 46: 115–124 - PubMed
-
- Aguilera A, Garcia-Muse T 2013. Causes of genome instability. Annu Rev Genet 47: 1–32 - PubMed
-
- Aguilera A, Gomez-Gonzalez B 2008. Genome instability: A mechanistic view of its causes and consequences. Nat Rev Genet 9: 204–217 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources