

Interplay between sirtuins, MYC and hypoxia-inducible factor in cancer-associated metabolic reprogramming
- PMID: 25085992
- PMCID: PMC4142723
- DOI: 10.1242/dmm.016287
Interplay between sirtuins, MYC and hypoxia-inducible factor in cancer-associated metabolic reprogramming
Abstract
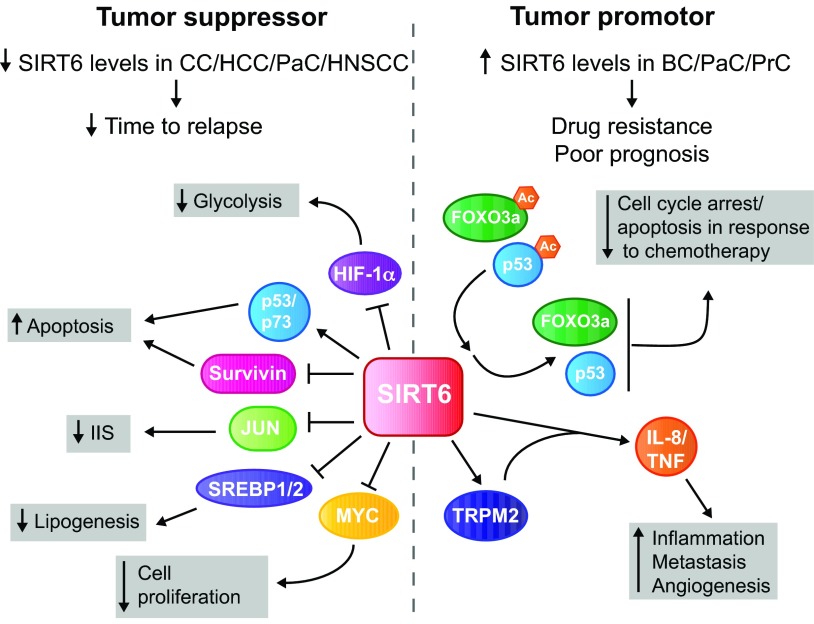
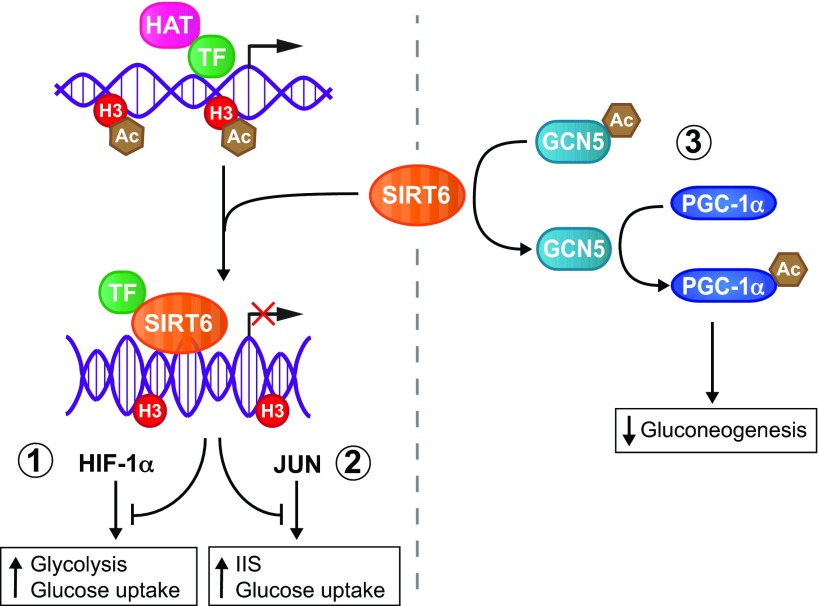
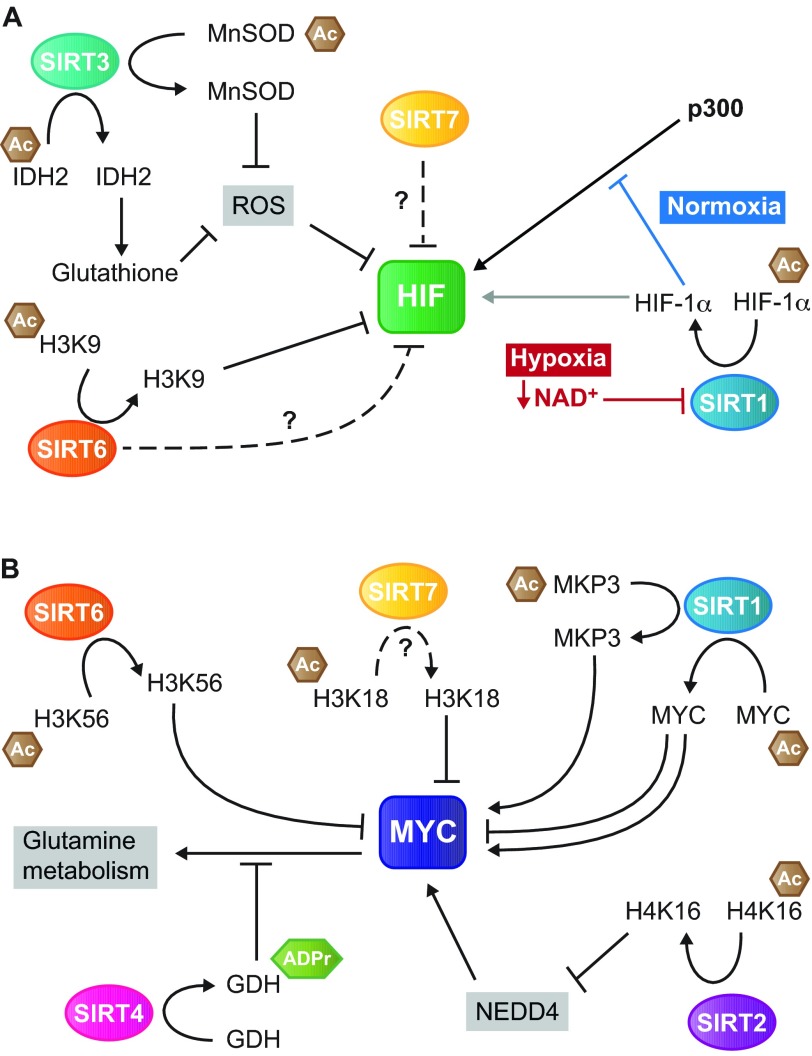
In the early twentieth century, Otto Heinrich Warburg described an elevated rate of glycolysis occurring in cancer cells, even in the presence of atmospheric oxygen (the Warburg effect). Despite the inefficiency of ATP generation through glycolysis, the breakdown of glucose into lactate provides cancer cells with a number of advantages, including the ability to withstand fluctuations in oxygen levels, and the production of intermediates that serve as building blocks to support rapid proliferation. Recent evidence from many cancer types supports the notion that pervasive metabolic reprogramming in cancer and stromal cells is a crucial feature of neoplastic transformation. Two key transcription factors that play major roles in this metabolic reprogramming are hypoxia inducible factor-1 (HIF1) and MYC. Sirtuin-family deacetylases regulate diverse biological processes, including many aspects of tumor biology. Recently, the sirtuin SIRT6 has been shown to inhibit the transcriptional output of both HIF1 and MYC, and to function as a tumor suppressor. In this Review, we highlight the importance of HIF1 and MYC in regulating tumor metabolism and their regulation by sirtuins, with a main focus on SIRT6.
Keywords: HIF; MYC; Metabolic reprogramming; Sirtuins; Warburg effect.
© 2014. Published by The Company of Biologists Ltd.
Figures
References
-
- Ahuja N., Schwer B., Carobbio S., Waltregny D., North B. J., Castronovo V., Maechler P., Verdin E. (2007). Regulation of insulin secretion by SIRT4, a mitochondrial ADP-ribosyltransferase. J. Biol. Chem. 282, 33583–33592 - PubMed
-
- Aury-Landas J., Bougeard G., Castel H., Hernandez-Vargas H., Drouet A., Latouche J. B., Schouft M. T., Férec C., Leroux D., Lasset C., et al. (2013). Germline copy number variation of genes involved in chromatin remodelling in families suggestive of Li-Fraumeni syndrome with brain tumours. Eur. J. Hum. Genet. 21, 1369–1376 - PMC - PubMed
-
- Bauer I., Grozio A., Lasigliè D., Basile G., Sturla L., Magnone M., Sociali G., Soncini D., Caffa I., Poggi A., et al. (2012). The NAD+-dependent histone deacetylase SIRT6 promotes cytokine production and migration in pancreatic cancer cells by regulating Ca2+ responses. J. Biol. Chem. 287, 40924–40937 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
