

Clinical and biochemical function of polymorphic NR0B1 GGAA-microsatellites in Ewing sarcoma: a report from the Children's Oncology Group
- PMID: 25093581
- PMCID: PMC4122435
- DOI: 10.1371/journal.pone.0104378
Clinical and biochemical function of polymorphic NR0B1 GGAA-microsatellites in Ewing sarcoma: a report from the Children's Oncology Group
Abstract
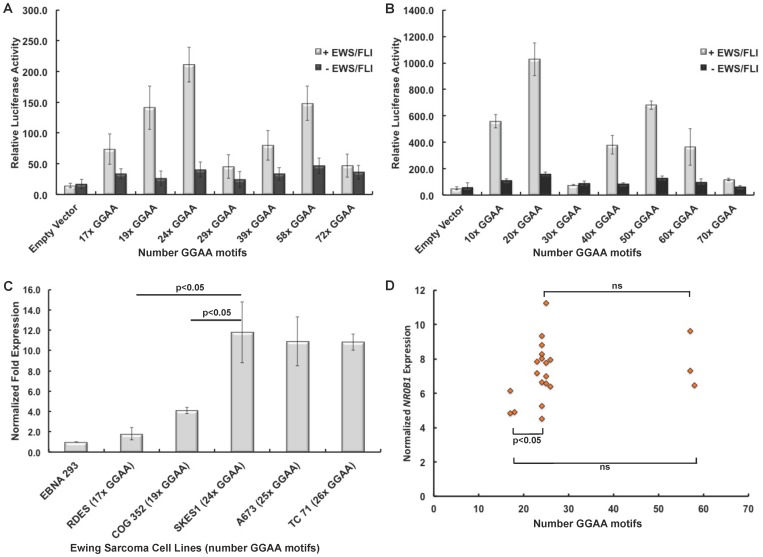
Background: The genetics involved in Ewing sarcoma susceptibility and prognosis are poorly understood. EWS/FLI and related EWS/ETS chimeras upregulate numerous gene targets via promoter-based GGAA-microsatellite response elements. These microsatellites are highly polymorphic in humans, and preliminary evidence suggests EWS/FLI-mediated gene expression is highly dependent on the number of GGAA motifs within the microsatellite.
Objectives: Here we sought to examine the polymorphic spectrum of a GGAA-microsatellite within the NR0B1 promoter (a critical EWS/FLI target) in primary Ewing sarcoma tumors, and characterize how this polymorphism influences gene expression and clinical outcomes.
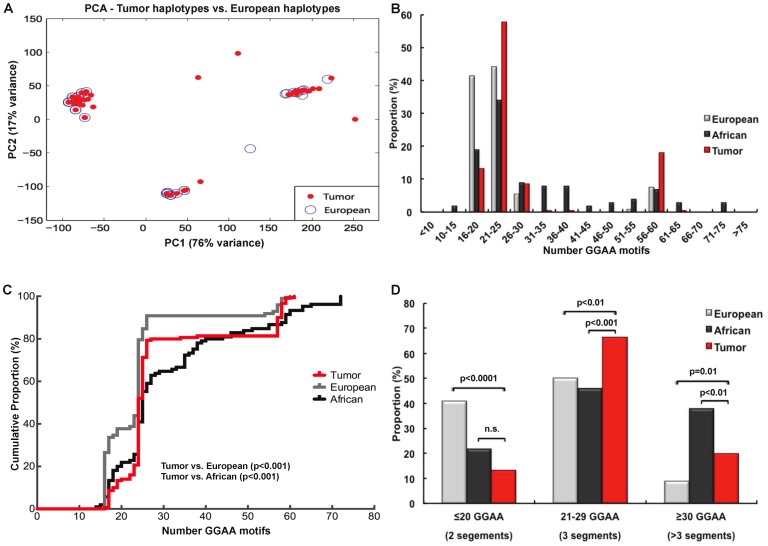
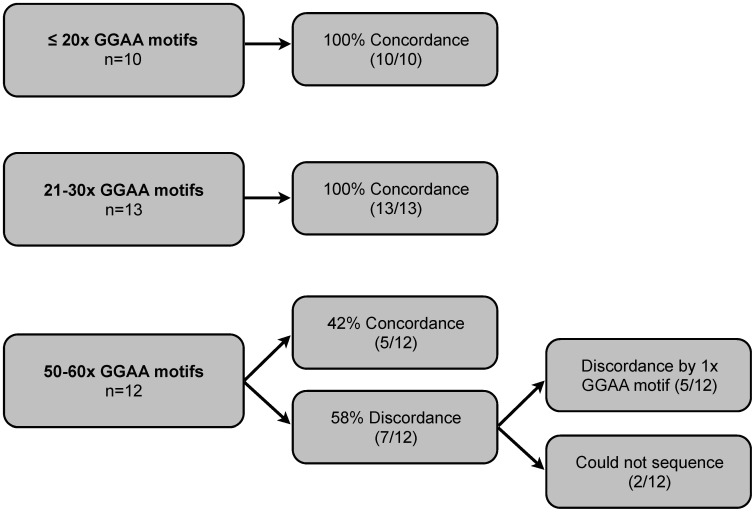
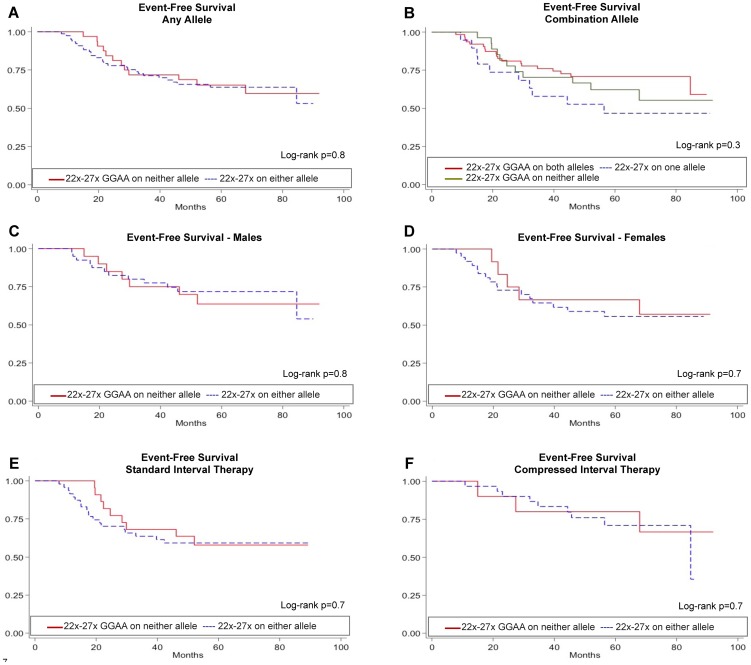
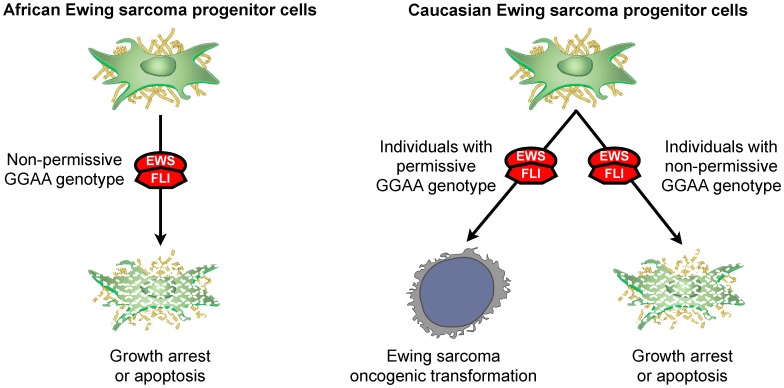
Results: A complex, bimodal pattern of EWS/FLI-mediated gene expression was observed across a wide range of GGAA motifs, with maximal expression observed in constructs containing 20-26 GGAA motifs. Relative to white European and African controls, the NR0B1 GGAA-microsatellite in tumor cells demonstrated a strong bias for haplotypes containing 21-25 GGAA motifs suggesting a relationship between microsatellite function and disease susceptibility. This selection bias was not a product of microsatellite instability in tumor samples, nor was there a correlation between NR0B1 GGAA-microsatellite polymorphisms and survival outcomes.
Conclusions: These data suggest that GGAA-microsatellite polymorphisms observed in human populations modulate EWS/FLI-mediated gene expression and may influence disease susceptibility in Ewing sarcoma.
Conflict of interest statement
Figures
References
-
- Delattre O, Zucman J, Melot T, Garau XS, Zucker JM, et al. (1994) The Ewing family of tumors–a subgroup of small-round-cell tumors defined by specific chimeric transcripts. N Engl J Med 331: 294–299. - PubMed
-
- Smith R, Owen LA, Trem DJ, Wong JS, Whangbo JS, et al. (2006) Expression profiling of EWS/FLI identifies NKX2.2 as a critical target gene in Ewing's sarcoma. Cancer cell 9: 405–416. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
