

Photobiont selectivity leads to ecological tolerance and evolutionary divergence in a polymorphic complex of lichenized fungi
- PMID: 25096324
- PMCID: PMC4204673
- DOI: 10.1093/aob/mcu146
Photobiont selectivity leads to ecological tolerance and evolutionary divergence in a polymorphic complex of lichenized fungi
Abstract
Background and aims: The integrity and evolution of lichen symbioses depend on a fine-tuned combination of algal and fungal genotypes. Geographically widespread species complexes of lichenized fungi can occur in habitats with slightly varying ecological conditions, and it remains unclear how this variation correlates with symbiont selectivity patterns in lichens. In an attempt to address this question, >300 samples were taken of the globally distributed and ecologically variable lichen-forming species complex Tephromela atra, together with closely allied species, in order to study genetic diversity and the selectivity patterns of their photobionts.
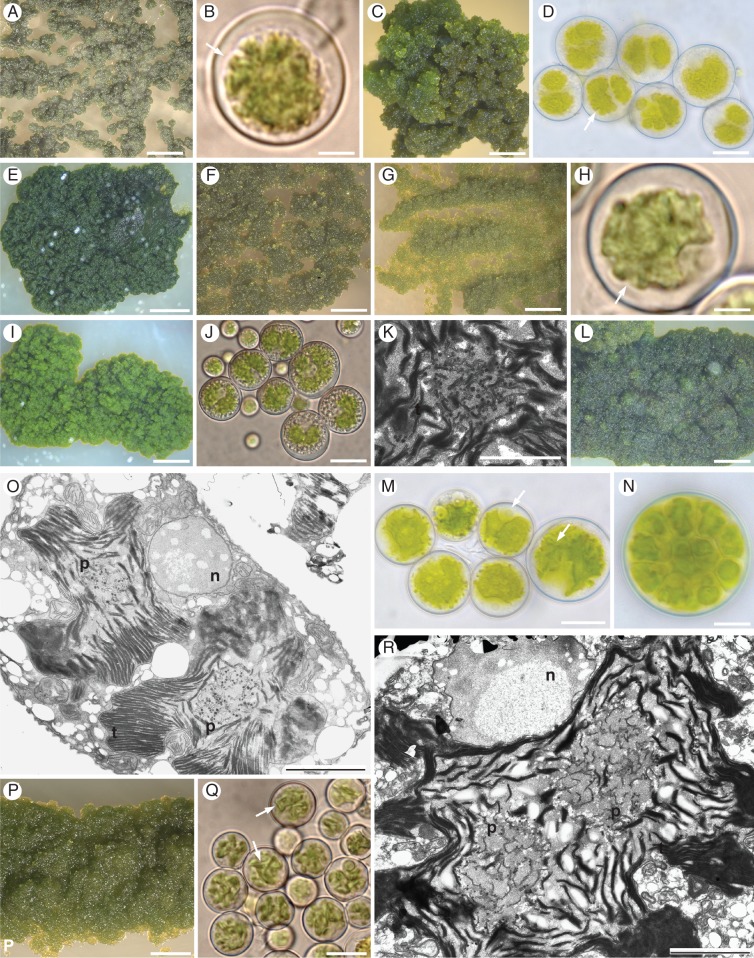
Methods: Lichen thalli of T. atra and of closely related species T. grumosa, T. nashii and T. atrocaesia were collected from six continents, across 24 countries and 62 localities representing a wide range of habitats. Analyses of genetic diversity and phylogenetic relationships were carried out both for photobionts amplified directly from the lichen thalli and from those isolated in axenic cultures. Morphological and anatomical traits were studied with light and transmission electron microscopy in the isolated algal strains.
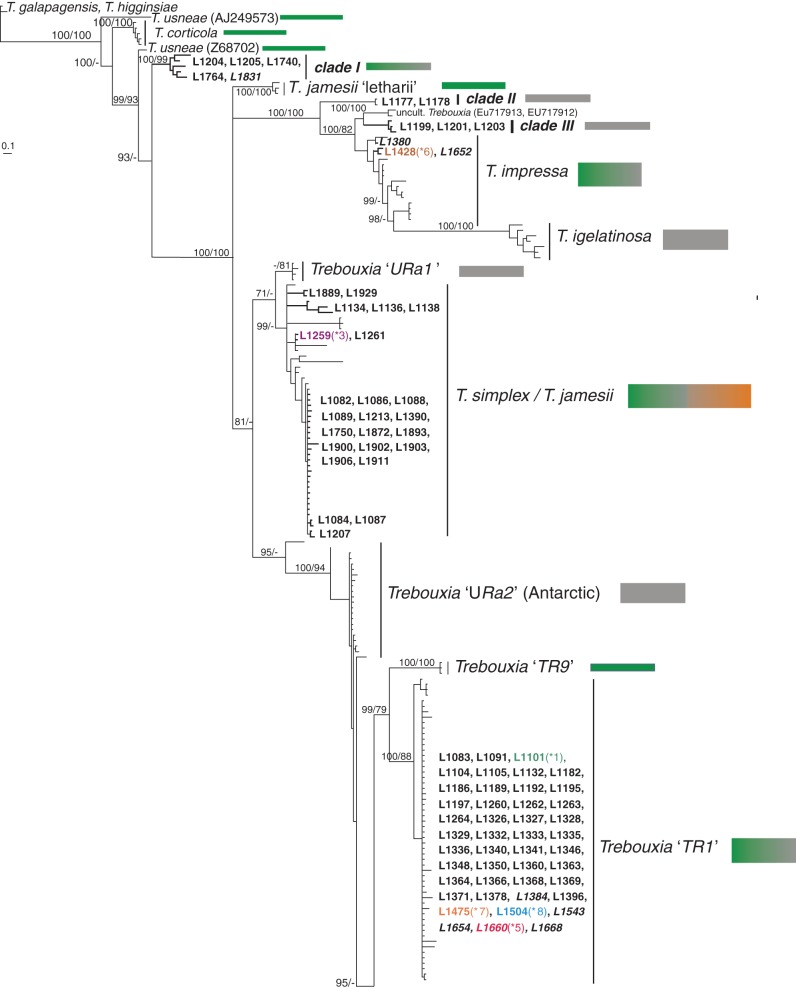
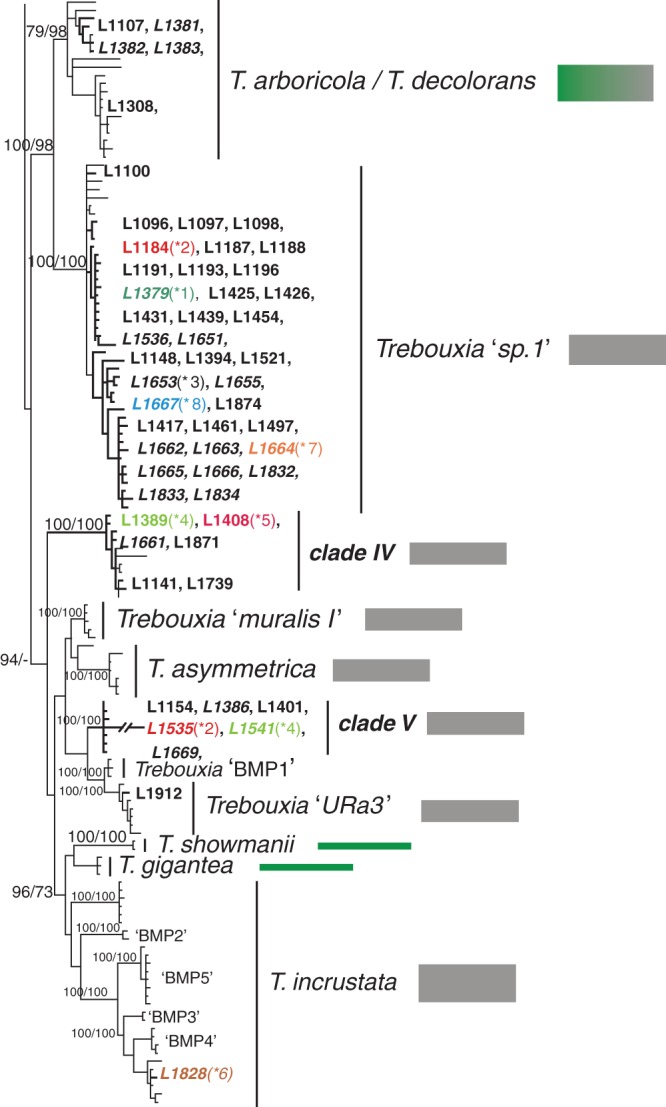
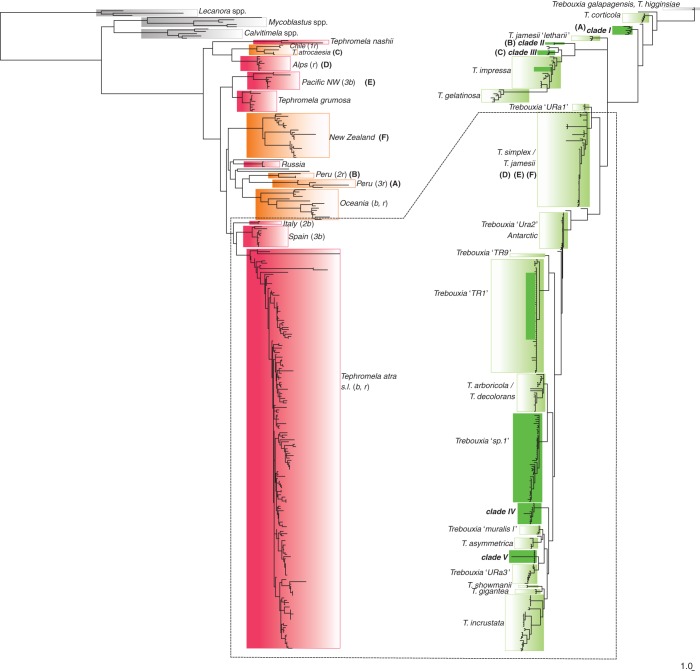
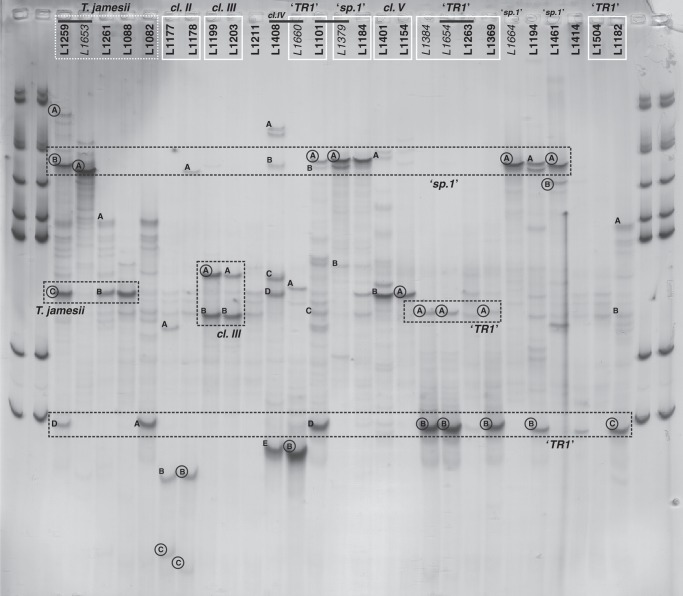
Key results: Tephromela fungal species were found to associate with 12 lineages of Trebouxia. Five new clades demonstrate the still-unrecognized genetic diversity of lichen algae. Culturable, undescribed lineages were also characterized by phenotypic traits. Strong selectivity of the mycobionts for the photobionts was observed in six monophyletic Tephromela clades. Seven Trebouxia lineages were detected in the poorly resolved lineage T. atra sensu lato, where co-occurrence of multiple photobiont lineages in single thalli was repeatedly observed.
Conclusions: Low selectivity apparently allows widespread lichen-forming fungi to establish successful symbioses with locally adapted photobionts in a broader range of habitats. This flexibility might correlate with both lower phylogenetic resolution and evolutionary divergence in species complexes of crustose lichen-forming fungi.
Keywords: Adaptation; Lecanoromycetes; Tephromela atra; Trebouxia; algal culture; crustose lichen; lichenized fungi; morphology; mycobiont; photobiont; phylogeny.
© The Author 2014. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Ahmadjian V. A guide to the algae occurring as lichen symbionts: isolation, culture, cultural physiology, and identification. Phycologia. 1967;6:127–160.
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. Journal of Molecular Biology. 1990;215:403–410. - PubMed
-
- Alvarez R, del Hoyo A, García-Breijo F, et al. Different strategies to achieve Pb-tolerance by the two Trebouxia algae coexisting in the lichen Ramalina farinacea. Journal of Plant Physiology. 2012;169:1797–806. - PubMed
-
- Beaumont HJE, Gallie J, Kost C, Ferguson GC, Rainey PB. Experimental evolution of bet hedging. Nature Letters. 2009;462:90–93. - PubMed
-
- Beck A. Photobionts: diversity and selectivity in lichen symbioses. International Lichenological Newsletter. 2002;35:18–24.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
