

Promiscuous methionyl-tRNA synthetase mediates adaptive mistranslation to protect cells against oxidative stress
- PMID: 25097229
- PMCID: PMC4179492
- DOI: 10.1242/jcs.152470
Promiscuous methionyl-tRNA synthetase mediates adaptive mistranslation to protect cells against oxidative stress
Abstract
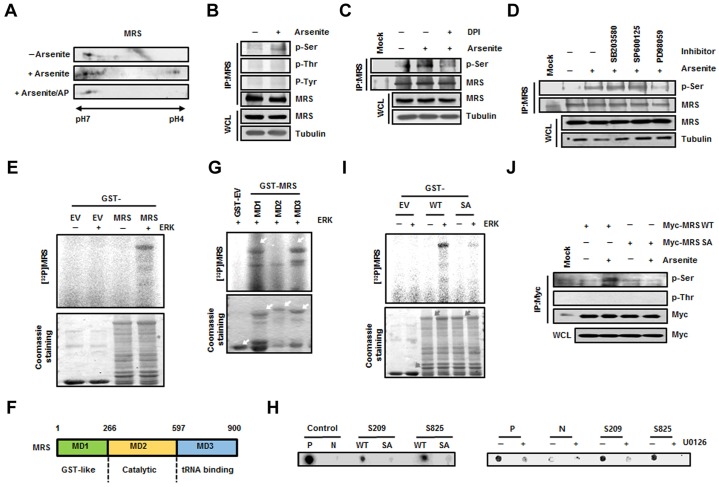
Aminoacyl-tRNA synthetases (ARSs) acylate transfer (t)RNAs with amino acids. Charging tRNAs with the right amino acids is the first step in translation; therefore, the accurate and error-free functioning of ARSs is an essential prerequisite for translational fidelity. A recent study found that methionine (Met) can be incorporated into non-Met residues of proteins through methionylation of non-cognate tRNAs under conditions of oxidative stress. However, it was not understood how this mis-methionylation is achieved. Here, we report that methionyl-tRNA synthetase (MRS) is phosphorylated at Ser209 and Ser825 by extracellular signal-related kinase (ERK1/2) under conditions of stress caused by reactive oxygen species (ROS), and that this phosphorylated MRS shows increased affinity for non-cognate tRNAs with lower affinity for tRNA(Met), leading to an increase in Met residues in cellular proteins. The expression of a mutant MRS containing the substitutions S209D and S825D, mimicking dual phosphorylation, reduced ROS levels and cell death. This controlled inaccuracy of MRS seems to serve as a defense mechanism against ROS-mediated damage at the cost of translational fidelity.
Keywords: Cell protection; ERK; Methionyl-tRNA synthetase; Misacylation; Reactive oxygen species.
© 2014. Published by The Company of Biologists Ltd.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
