

Manipulation of Fgf and Bmp signaling in teleost fishes suggests potential pathways for the evolutionary origin of multicuspid teeth
- PMID: 25098636
- PMCID: PMC4289623
- DOI: 10.1111/ede.12021
Manipulation of Fgf and Bmp signaling in teleost fishes suggests potential pathways for the evolutionary origin of multicuspid teeth
Abstract
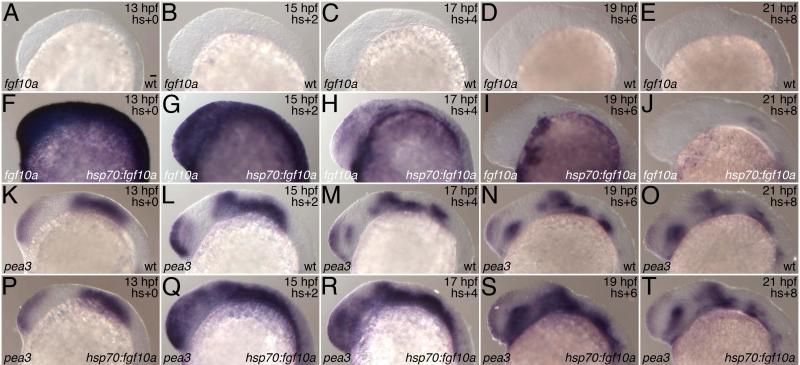
Teeth with two or more cusps have arisen independently from an ancestral unicuspid condition in a variety of vertebrate lineages, including sharks, teleost fishes, amphibians, lizards, and mammals. One potential explanation for the repeated origins of multicuspid teeth is the existence of multiple adaptive pathways leading to them, as suggested by their different uses in these lineages. Another is that the addition of cusps required only minor changes in genetic pathways regulating tooth development. Here we provide support for the latter hypothesis by demonstrating that manipulation of the levels of Fibroblast growth factor (Fgf) or Bone morphogenetic protein (Bmp) signaling produces bicuspid teeth in the zebrafish (Danio rerio), a species lacking multicuspid teeth in its ancestry. The generality of these results for teleosts is suggested by the conversion of unicuspid pharyngeal teeth into bicuspid teeth by similar manipulations of the Mexican Tetra (Astyanax mexicanus). That these manipulations also produced supernumerary teeth in both species supports previous suggestions of similarities in the molecular control of tooth and cusp number. We conclude that despite their apparent complexity, the evolutionary origin of multicuspid teeth is positively constrained, likely requiring only slight modifications of a pre-existing mechanism for patterning the number and spacing of individual teeth.
© 2013 Wiley Periodicals, Inc.
Figures






References
-
- Alexander RM. Adaptation in the skulls and cranial muscles of South American characinoid fish. J. Linn. Soc. Lond. Zool. 1964;45:169–190.
-
- Andreucci RD, Britski HA, Carneiro J. Structure and evolution of tetraodontoid teeth: an autoradiographic study (Pisces, Tetraodontiformes). J. Morphol. 1982;171:283–292. - PubMed
-
- Bolt JR. Lissamphibian origins. In: Schultze H-P, Trueb L, editors. Origins of the Higher Groups of Tetrapods. Cornell University Press; Ithaca, NY: 1991. pp. 194–220.
-
- Carroll RL. Vertebrate Paleontology and Evolution. W. H. Freeman; New York: 1988.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
