

Characterization of a novel water pocket inside the human Cx26 hemichannel structure
- PMID: 25099799
- PMCID: PMC4129493
- DOI: 10.1016/j.bpj.2014.05.037
Characterization of a novel water pocket inside the human Cx26 hemichannel structure
Abstract
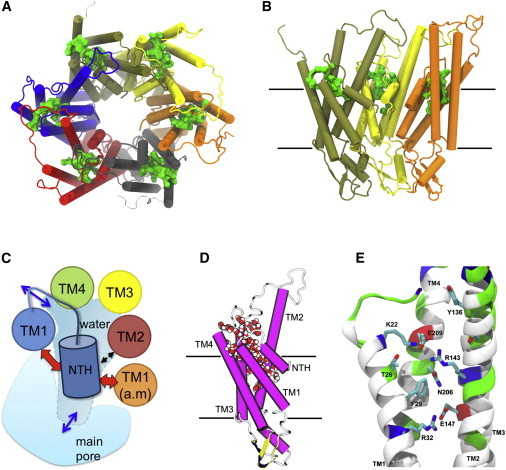
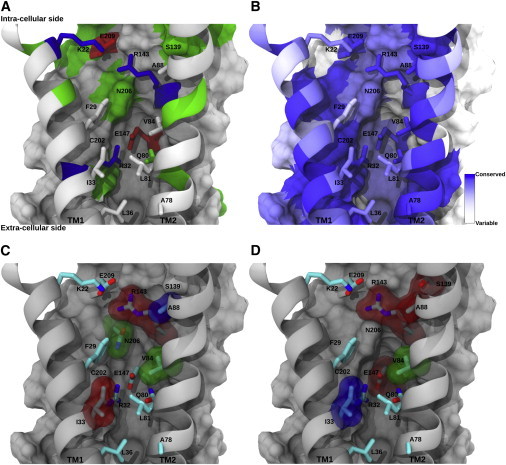
Connexins (Cxs) are a family of vertebrate proteins constituents of gap junction channels (GJCs) that connect the cytoplasm of adjacent cells by the end-to-end docking of two Cx hemichannels. The intercellular transfer through GJCs occurs by passive diffusion allowing the exchange of water, ions, and small molecules. Despite the broad interest to understand, at the molecular level, the functional state of Cx-based channels, there are still many unanswered questions regarding structure-function relationships, perm-selectivity, and gating mechanisms. In particular, the ordering, structure, and dynamics of water inside Cx GJCs and hemichannels remains largely unexplored. In this work, we describe the identification and characterization of a believed novel water pocket-termed the IC pocket-located in-between the four transmembrane helices of each human Cx26 (hCx26) monomer at the intracellular (IC) side. Using molecular dynamics (MD) simulations to characterize hCx26 internal water structure and dynamics, six IC pockets were identified per hemichannel. A detailed characterization of the dynamics and ordering of water including conformational variability of residues forming the IC pockets, together with multiple sequence alignments, allowed us to propose a functional role for this cavity. An in vitro assessment of tracer uptake suggests that the IC pocket residue Arg-143 plays an essential role on the modulation of the hCx26 hemichannel permeability.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Jensen L.H. The structure of water in protein crystals. Dev. Biol. Stand. 1992;74:53–61. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
