

Review
doi: 10.1038/nm.3643.
Emerging interactions between skin stem cells and their niches
Affiliations
- PMID: 25100530
- PMCID: PMC4358898
- DOI: 10.1038/nm.3643
Item in Clipboard
Review
Emerging interactions between skin stem cells and their niches
Nat Med.
2014 Aug.
Abstract
The skin protects mammals from insults, infection and dehydration and enables thermoregulation and sensory perception. Various skin-resident cells carry out these diverse functions. Constant turnover of cells and healing upon injury necessitate multiple reservoirs of stem cells. Thus, the skin provides a model for studying interactions between stem cells and their microenvironments, or niches. Advances in genetic and imaging tools have brought new findings about the lineage relationships between skin stem cells and their progeny and about the mutual influences between skin stem cells and their niches. Such knowledge may offer novel avenues for therapeutics and regenerative medicine.
Figures
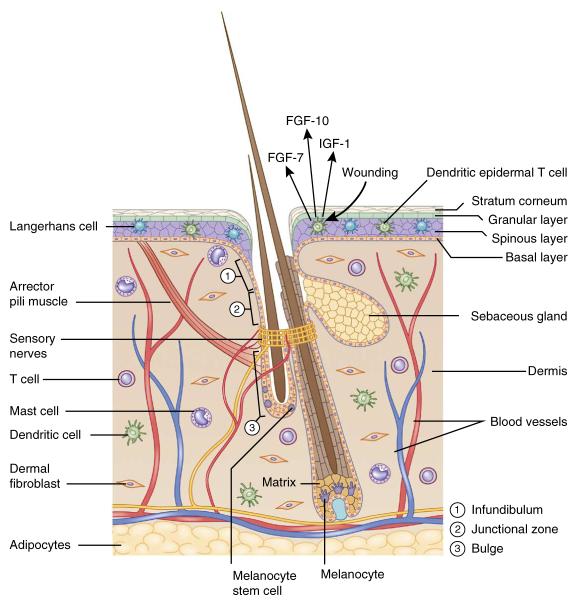
The skin: an organ with a diverse array of cell types. The hair follicle is a complex appendage of the epidermis. It is composed of an infundibulum that opens to the skin surface, sebaceous glands, and the junctional zone between the glands and the bulge. Hair follicle and melanocyte stem cells reside in the bulge and the hair germ. In full anagen, hair follicle stem cells regenerate the lower two-thirds of the follicle, including the matrix, which produces the hair and its channel. Melanocyte stem cells generate mature melanocytes, which transfer their pigment to differentiating hair cells. The hair follicle also serves as a hub attracting peripheral nerves, blood vessels and arrector pili muscles. The dermis is populated with dermal fibroblasts and various immune cells such as mast cells, dendritic cells and T cells. Deeper in the dermis is a layer of subcutaneous adipocytes.
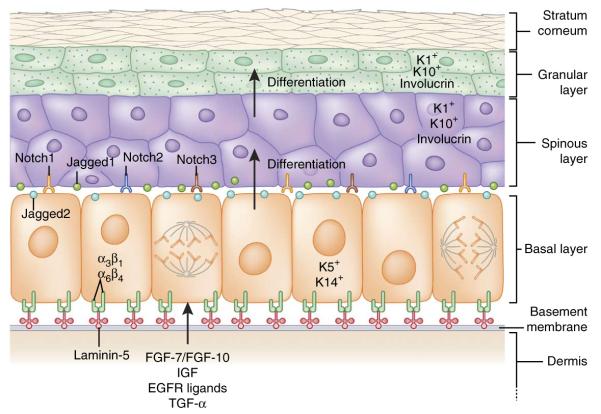
Interfollicular epidermis: architecture, signaling and lineages. The epidermis is a stratified structure. Self-renewing stem cells reside within the basal layer, which adheres through α3β1 and α6β4 integrins to an underlying basement membrane of laminin-5–rich extracellular matrix that separates the epidermis from the underlying dermis. Secreted factors such as FGF-7, FGF-10, IGF, EGF ligands and TGF-α from dermal fibroblasts promote the proliferation of basal epidermal cells. Proliferative basal progenitors generate columnar units of Notch-activated terminally differentiating cells that go through three stages: spinous layers, granular layers and finally dead stratum corneum layers that eventually are shed from the skin surface. Each cell type expresses a different gamut of keratin (K) proteins.
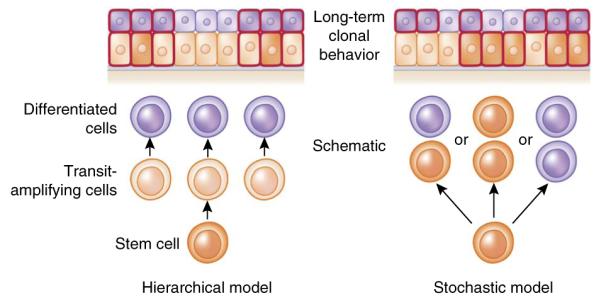
Hierarchical versus stochastic models of epidermal differentiation. In a hierarchical model, rare divisions by stem cells generate rapidly dividing transit amplifying cells, which then give rise to differentiated cells. During lineage tracing, only clones marking the stem cells are long lived, and thus clone sizes become invariant after a period of time. By contrast, in a stochastic model, all basal cells are the same and each division can yield three different outcomes: (i) one differentiated daughter that withdraws from cell cycle and departs from the basal layer, and one progenitor that remains in the basal layer and continues to divide; (ii) two basal progenitors; and (iii) two differentiated daughters. Although the fate choices are random, the probabilities of different outcomes are similar, so that the generation of differentiated cells and the maintenance of committed progenitor pools are balanced at the population level and long-term homeostasis is ensured. In this model, each individual clone will vary in size. Predictions of lineage-tracing results from each model are shown at the top of the diagram; cells outlined in red are the ones retaining lineage-traced marks.
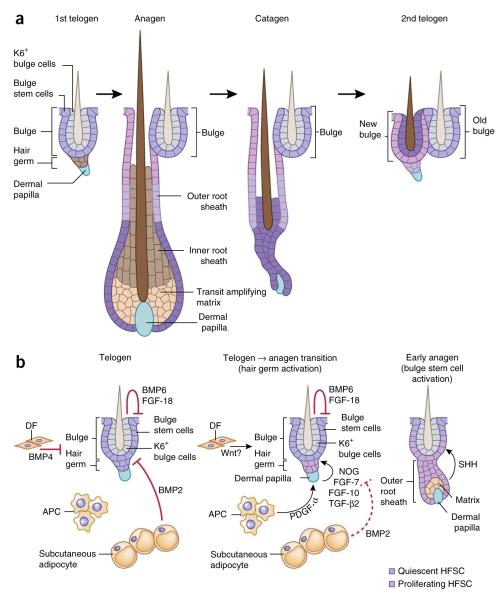
Hair follicle lineage and niche signals regulate hair follicle stem cells. (a) HFSCs can exist in two states. Quiescent bulge stem cells (Bu-SCs) are located in the outer layer of this niche and contribute to the generation of the outer root sheath. Primed stem cells reside in the hair germ, sandwiched between the bulge and a specialized dermal cluster known as the dermal papilla. They are responsible for generating the transit amplifying cell (TAC) matrix, which then gives rise to the hair shaft and its inner root sheath (IRS) channel. Although matrix and IRS are destroyed during catagen, many of the outer root sheath (ORS) cells are spared and generate a new bulge right next to the original one at the end of catagen. The upper ORS contributes to the outer layer of the new bulge, and the middle ORS contributes to the hair germ. Some of the lower ORS cells become the differentiated inner keratin 6+ (K6+) bulge cells, which provide inhibitory signals to Bu-SCs, raising their activation threshold for the next hair cycle. (b) During telogen, K6+ bulge cells produce BMP6 and FGF-18, dermal fibroblasts (DFs) produce BMP4 and subcutaneous adipocytes express BMP2. Together, these factors maintain Bu-SCs and hair germ in quiescence. At the transition to anagen, BMP2 and BMP4 are downregulated, whereas the expression of activation factors including noggin (NOG), FGF-7, FGF-10 and TGF-β2 from dermal papillae and PDGF-α from adipocyte precursor cells (APCs) is elevated. This, in turn, stimulates hair germ proliferation, and a new hair cycle is launched. Bu-SCs maintain their quiescent state until TAC matrix is generated and starts producing SHH.
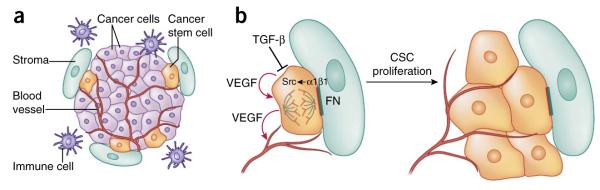
Signaling pathways in skin cancers. (a) The squamous cell carcinoma–cancer stem cell niche. CSCs are often found at the tumor-stroma interface, together with an elaborate vasculature, immune cells and aberrant fibroblasts. (b) Upon activation of αβ1 integrins by extracellular matrix ligands such as fibronectin (FN), focal adhesion kinase (FAK) and its associate tyrosine kinase Src become hyperactivated and promote proliferation of CSCs. By contrast, TGF-β signaling counteracts integrin activity and enhances CSC quiescence. In addition, CSCs secrete VEGF, which acts in an autocrine fashion to enhance CSC proliferation and in a paracrine fashion to promote formation of new blood vessels.
References
-
- Xie T, Spradling AC. A niche maintaining germ line stem cells in the Drosophila ovary. Science. 2000;290:328–330. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
