

Conformational analysis of the Streptococcus pneumoniae hyaluronate lyase and characterization of its hyaluronan-specific carbohydrate-binding module
- PMID: 25100731
- PMCID: PMC4175358
- DOI: 10.1074/jbc.M114.578435
Conformational analysis of the Streptococcus pneumoniae hyaluronate lyase and characterization of its hyaluronan-specific carbohydrate-binding module
Abstract
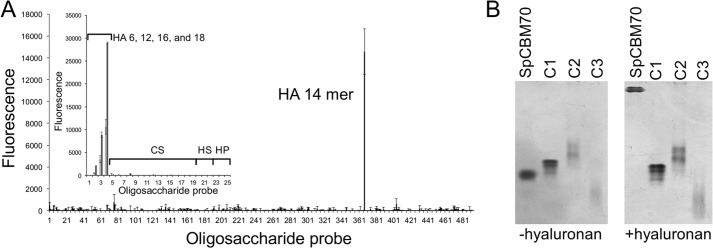
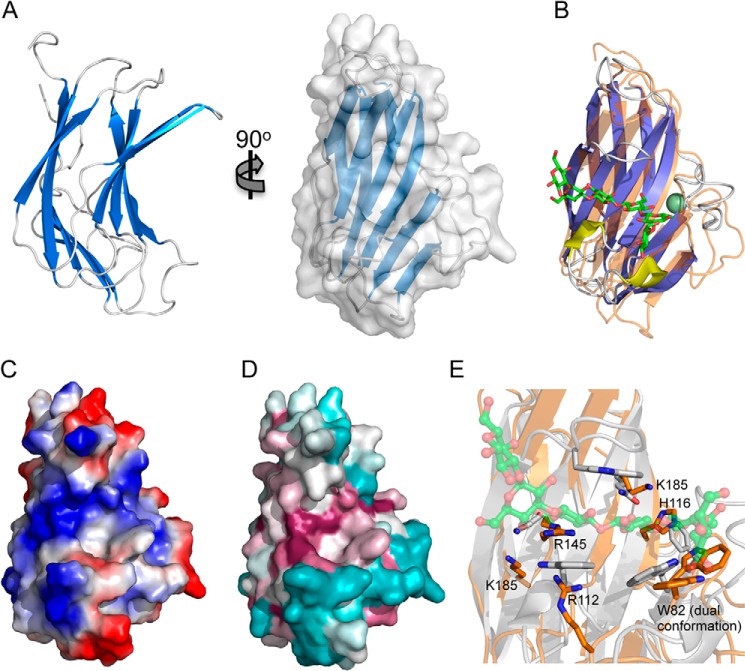
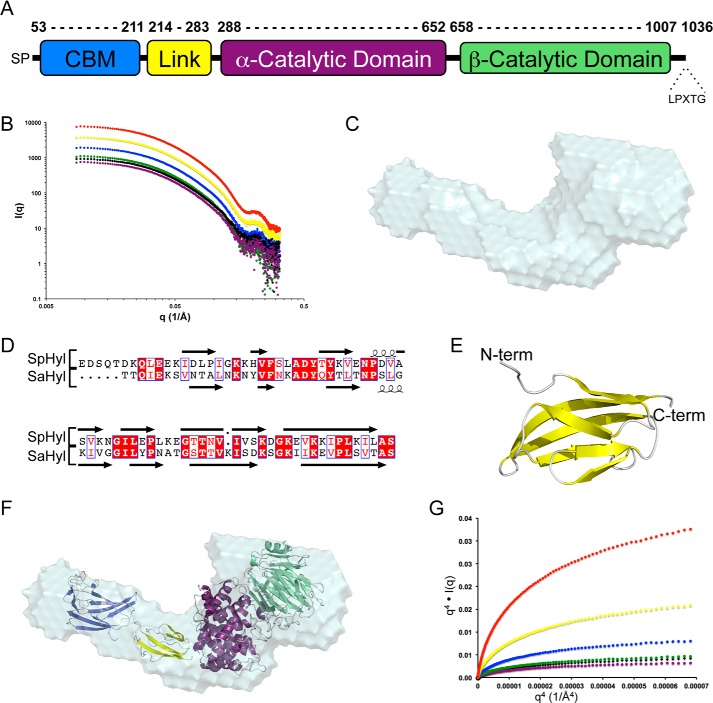
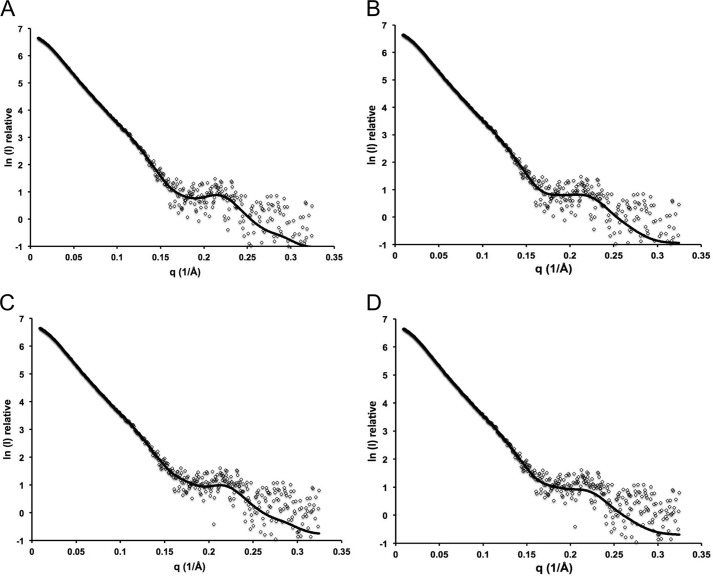
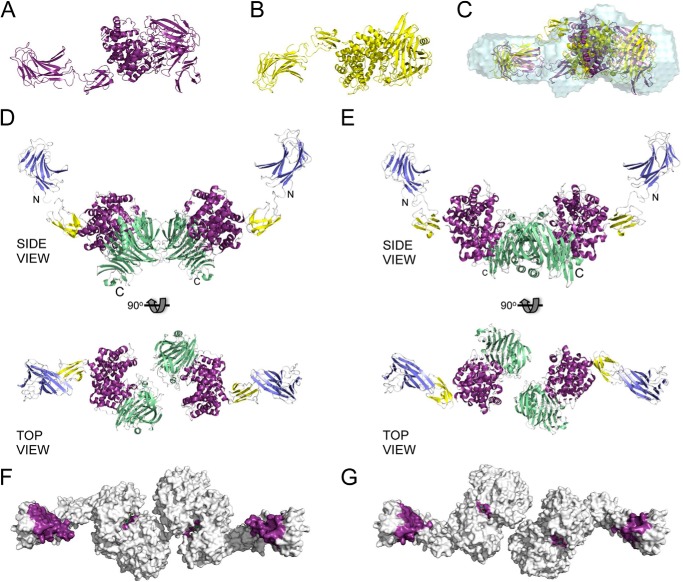
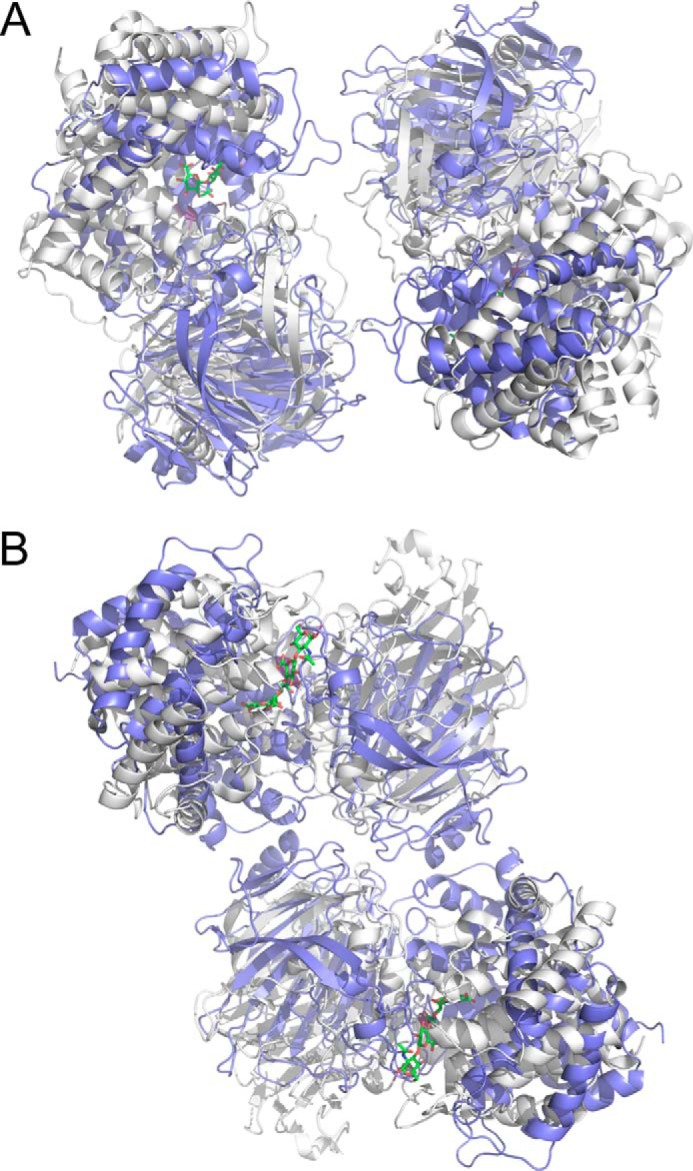
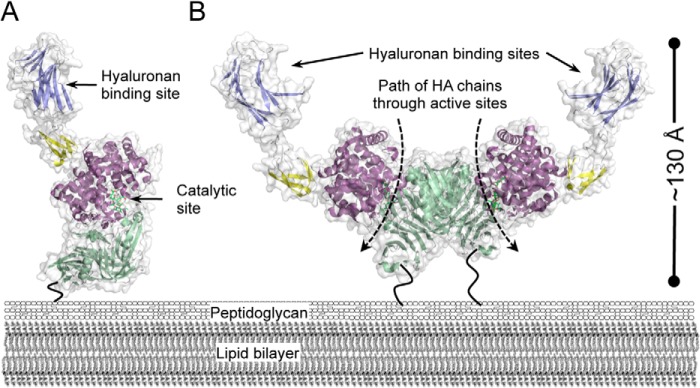
For a subset of pathogenic microorganisms, including Streptococcus pneumoniae, the recognition and degradation of host hyaluronan contributes to bacterial spreading through the extracellular matrix and enhancing access to host cell surfaces. The hyaluronate lyase (Hyl) presented on the surface of S. pneumoniae performs this role. Using glycan microarray screening, affinity electrophoresis, and isothermal titration calorimetry we show that the N-terminal module of Hyl is a hyaluronan-specific carbohydrate-binding module (CBM) and the founding member of CBM family 70. The 1.2 Å resolution x-ray crystal structure of CBM70 revealed it to have a β-sandwich fold, similar to other CBMs. The electrostatic properties of the binding site, which was identified by site-directed mutagenesis, are distinct from other CBMs and complementary to its acidic ligand, hyaluronan. Dynamic light scattering and solution small angle x-ray scattering revealed the full-length Hyl protein to exist as a monomer/dimer mixture in solution. Through a detailed analysis of the small angle x-ray scattering data, we report the pseudoatomic solution structures of the monomer and dimer forms of the full-length multimodular Hyl.
Keywords: Carbohydrate; Carbohydrate Processing; Carbohydrate-binding Protein; Crystallography; Glycobiology; Hyaluronan; Hyaluronate Lyase; Small Angle X-ray Scattering (SAXS); Streptococcus.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Rapport M. M., Linker A., Meyer K. (1951) The hydrolysis of hyaluronic acid by pneumococcal hyaluronidase. J. Biol. Chem. 192, 283–291 - PubMed
-
- Thompson R. T., Moses F. E. (1948) Elaboration of hyaluronidase by pneumococci isolated from bacteremic pneumococcic pneumonia patients. J. Clin. Invest. 27, 558. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
