

The 3' untranslated region of Pea Enation Mosaic Virus contains two T-shaped, ribosome-binding, cap-independent translation enhancers
- PMID: 25100834
- PMCID: PMC4178710
- DOI: 10.1128/JVI.01433-14
The 3' untranslated region of Pea Enation Mosaic Virus contains two T-shaped, ribosome-binding, cap-independent translation enhancers
Abstract
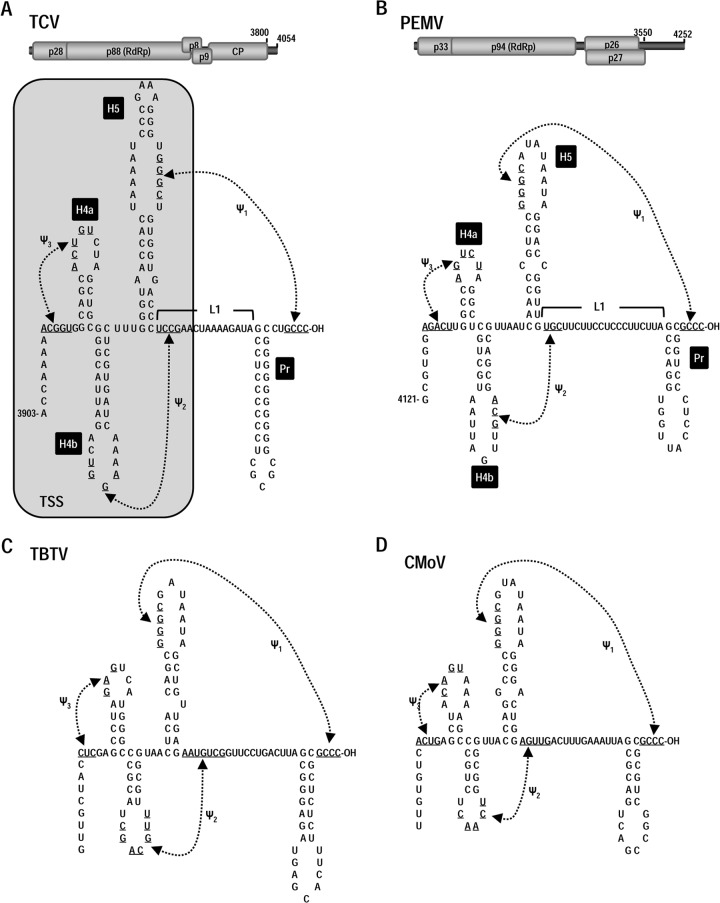
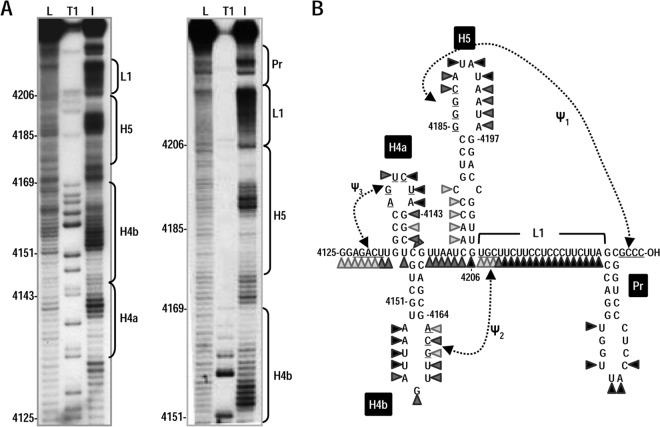
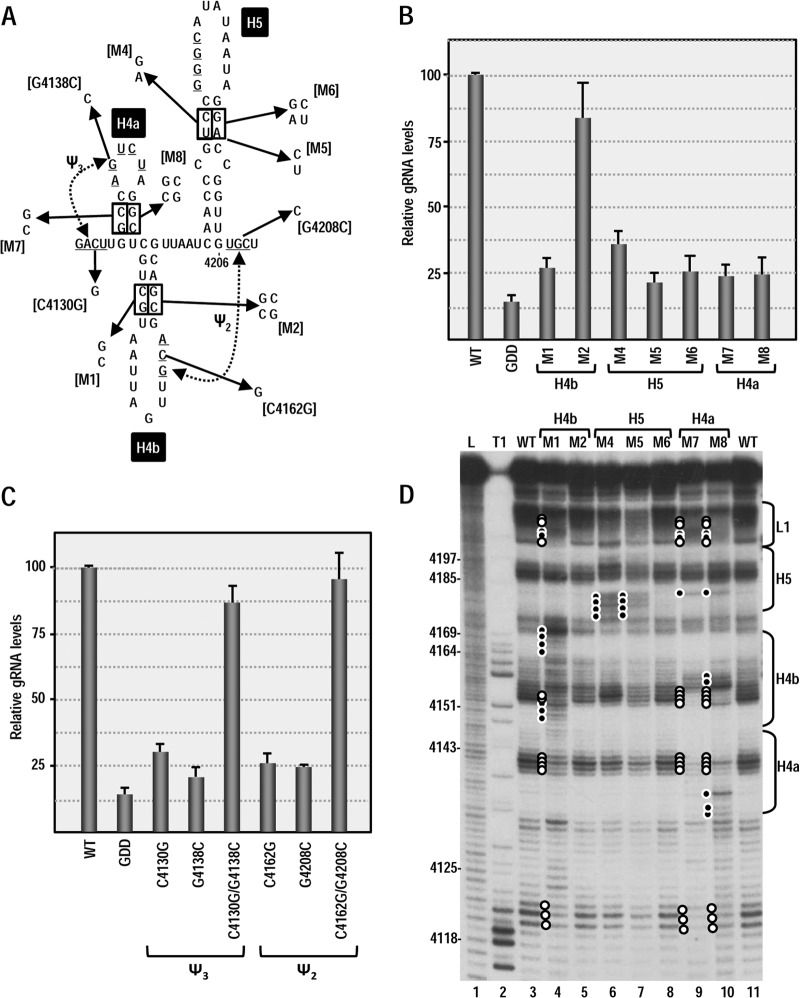
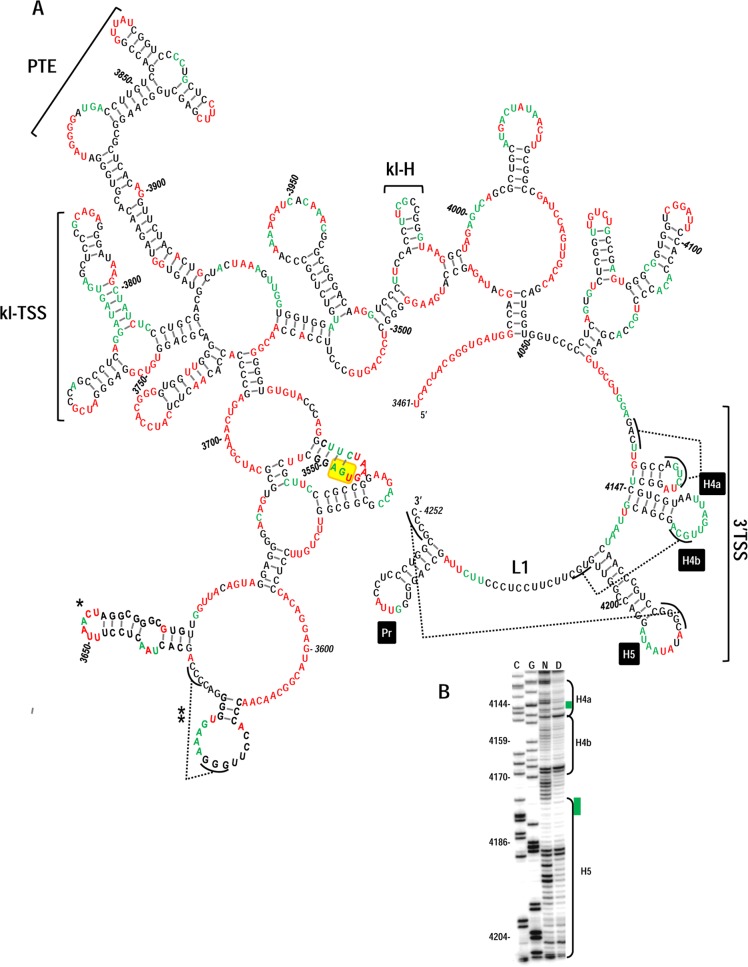
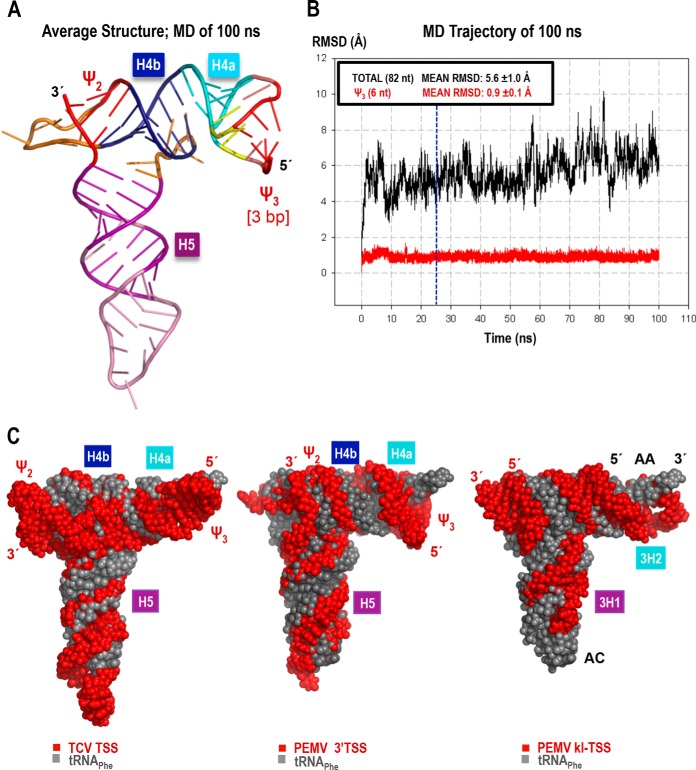
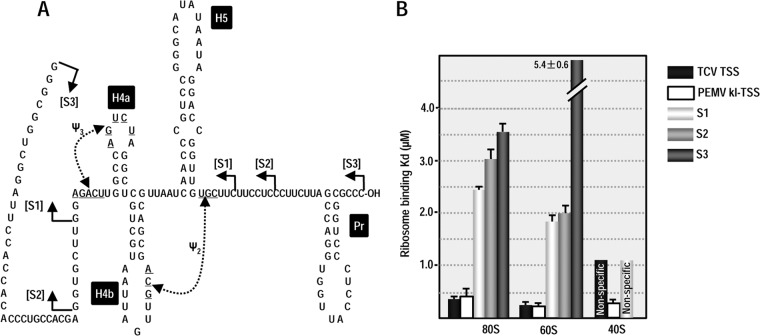
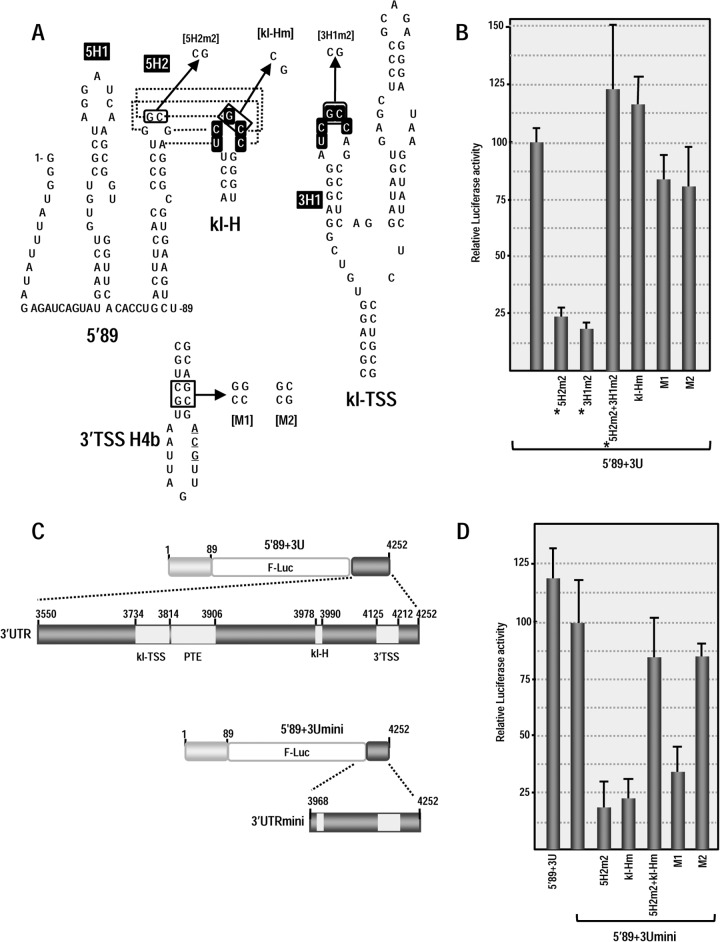
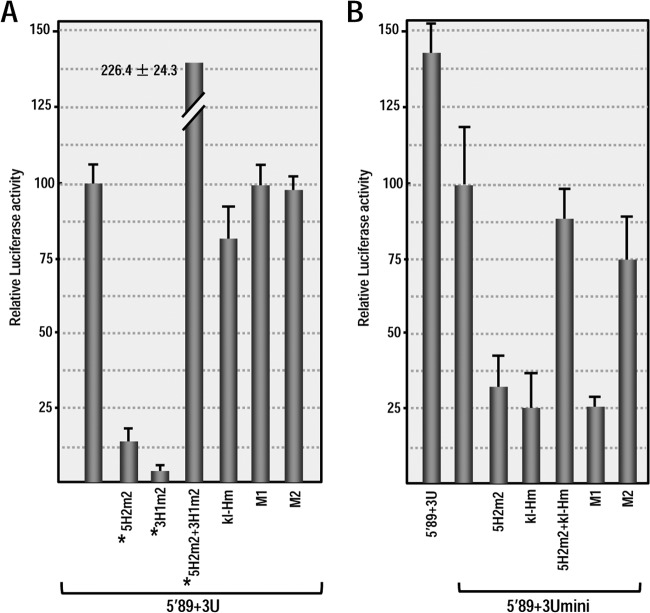
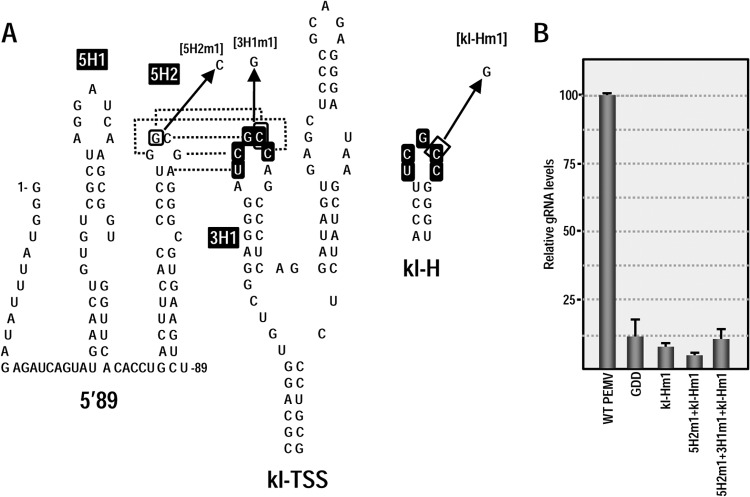
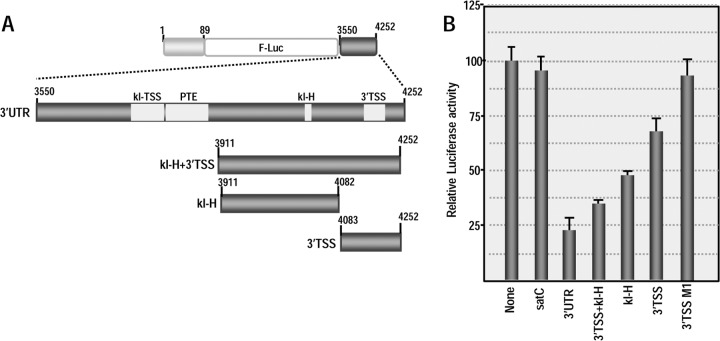
Many plant viruses without 5' caps or 3' poly(A) tails contain 3' proximal, cap-independent translation enhancers (3'CITEs) that bind to ribosomal subunits or translation factors thought to assist in ribosome recruitment. Most 3'CITEs participate in a long-distance kissing-loop interaction with a 5' proximal hairpin to deliver ribosomal subunits to the 5' end for translation initiation. Pea Enation Mosaic Virus (PEMV) contains two adjacent 3'CITEs in the center of its 703-nucleotide 3' untranslated region (3'UTR), the ribosome-binding, kissing-loop T-shaped structure (kl-TSS) and eukaryotic translation initiation factor 4E-binding Panicum mosaic virus-like translation enhance (PTE). We now report that PEMV contains a third, independent 3'CITE located near the 3' terminus. This 3'CITE is composed of three hairpins and two pseudoknots, similar to the TSS 3'CITE of the carmovirus Turnip crinkle virus (TCV). As with the TCV TSS, the PEMV 3'TSS is predicted to fold into a T-shaped structure that binds to 80S ribosomes and 60S ribosomal subunits. A small hairpin (kl-H) upstream of the 3'TSS contains an apical loop capable of forming a kissing-loop interaction with a 5' proximal hairpin and is critical for the accumulation of full-length PEMV in protoplasts. Although the kl-H and 3'TSS are dispensable for the translation of a reporter construct containing the complete PEMV 3'UTR in vitro, deleting the normally required kl-TSS and PTE 3'CITEs and placing the kl-H and 3'TSS proximal to the reporter termination codon restores translation to near wild-type levels. This suggests that PEMV requires three 3'CITEs for proper translation and that additional translation enhancers may have been missed if reporter constructs were used in 3'CITE identification. Importance: The rapid life cycle of viruses requires efficient translation of viral-encoded proteins. Many plant RNA viruses contain 3' cap-independent translation enhancers (3'CITEs) to effectively compete with ongoing host translation. Since only single 3'CITEs have been identified for the vast majority of individual viruses, it is widely accepted that this is sufficient for a virus's translational needs. Pea Enation Mosaic Virus possesses a ribosome-binding 3'CITE that can connect to the 5' end through an RNA-RNA interaction and an adjacent eukaryotic translation initiation factor 4E-binding 3'CITE. We report the identification of a third 3'CITE that binds weakly to ribosomes and requires an upstream hairpin to form a bridge between the 3' and 5' ends. Although both ribosome-binding 3'CITEs are critical for virus accumulation in vivo, only the CITE closest to the termination codon of a reporter open reading frame is active, suggesting that artificial constructs used for 3'CITE identification may underestimate the number of CITEs that participate in translation.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
