

Overcoming stalled translation in human mitochondria
- PMID: 25101074
- PMCID: PMC4103422
- DOI: 10.3389/fmicb.2014.00374
Overcoming stalled translation in human mitochondria
Abstract
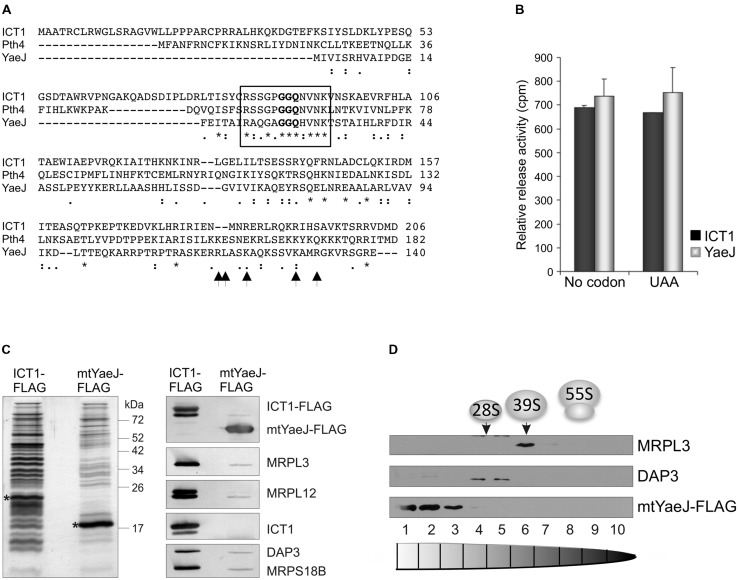
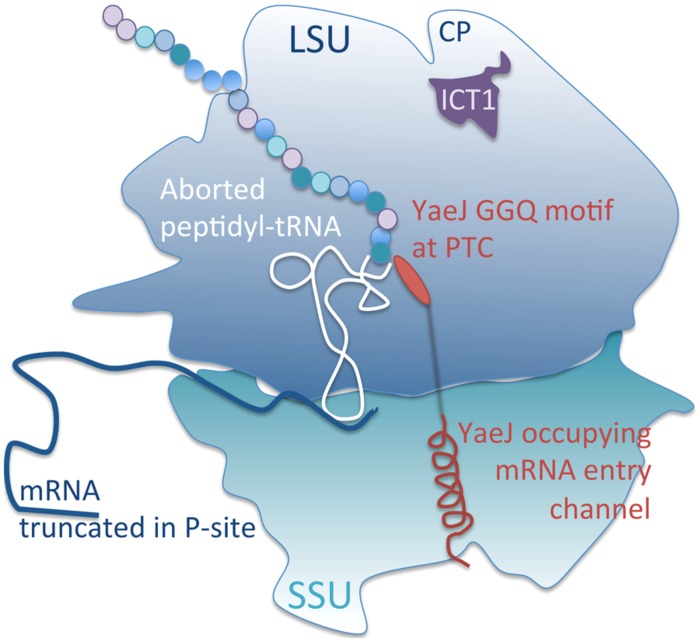
Protein synthesis is central to life and maintaining a highly accurate and efficient mechanism is essential. What happens when a translating ribosome stalls on a messenger RNA? Many highly intricate processes have been documented in the cytosol of numerous species, but how does organellar protein synthesis resolve this stalling issue? Mammalian mitochondria synthesize just thirteen highly hydrophobic polypeptides. These proteins are all integral components of the machinery that couples oxidative phosphorylation. Consequently, it is essential that stalled mitochondrial ribosomes can be efficiently recycled. To date, there is no evidence to support any particular molecular mechanism to resolve this problem. However, here we discuss the observation that there are four predicted members of the mitochondrial translation release factor family and that only one member, mtRF1a, is necessary to terminate the translation of all thirteen open reading frames in the mitochondrion. Could the other members be involved in the process of recycling stalled mitochondrial ribosomes?
Keywords: ICT1; mitochondria; protein synthesis; release factor; ribosome rescue; ribosome stalling; translation.
Figures
Similar articles
-
Ribosome rescue and translation termination at non-standard stop codons by ICT1 in mammalian mitochondria.PLoS Genet. 2014 Sep 18;10(9):e1004616. doi: 10.1371/journal.pgen.1004616. eCollection 2014 Sep. PLoS Genet. 2014. PMID: 25233460 Free PMC article.
-
Rescuing stalled mammalian mitoribosomes - what can we learn from bacteria?J Cell Sci. 2020 Jan 2;133(1):jcs231811. doi: 10.1242/jcs.231811. J Cell Sci. 2020. PMID: 31896602 Review.
-
A functional peptidyl-tRNA hydrolase, ICT1, has been recruited into the human mitochondrial ribosome.EMBO J. 2010 Mar 17;29(6):1116-25. doi: 10.1038/emboj.2010.14. Epub 2010 Feb 25. EMBO J. 2010. PMID: 20186120 Free PMC article.
-
Structural basis of translation termination, rescue, and recycling in mammalian mitochondria.Mol Cell. 2021 Jun 17;81(12):2566-2582.e6. doi: 10.1016/j.molcel.2021.03.042. Epub 2021 Apr 19. Mol Cell. 2021. PMID: 33878294
-
Maintaining mitochondrial ribosome function: The role of ribosome rescue and recycling factors.RNA Biol. 2022;19(1):117-131. doi: 10.1080/15476286.2021.2015561. Epub 2021 Dec 31. RNA Biol. 2022. PMID: 34923906 Free PMC article. Review.
Cited by
-
Mechanisms and regulation of protein synthesis in mitochondria.Nat Rev Mol Cell Biol. 2021 May;22(5):307-325. doi: 10.1038/s41580-021-00332-2. Epub 2021 Feb 16. Nat Rev Mol Cell Biol. 2021. PMID: 33594280 Review.
-
Dual function of GTPBP6 in biogenesis and recycling of human mitochondrial ribosomes.Nucleic Acids Res. 2020 Dec 16;48(22):12929-12942. doi: 10.1093/nar/gkaa1132. Nucleic Acids Res. 2020. PMID: 33264405 Free PMC article.
-
Knockdown of immature colon carcinoma transcript-1 inhibits proliferation of glioblastoma multiforme cells through Gap 2/mitotic phase arrest.Onco Targets Ther. 2015 May 19;8:1119-27. doi: 10.2147/OTT.S75864. eCollection 2015. Onco Targets Ther. 2015. Retraction in: Onco Targets Ther. 2023 Jun 27;16:479-480. doi: 10.2147/OTT.S427669. PMID: 26056476 Free PMC article. Retracted.
-
LISTERIN E3 Ubiquitin Ligase and Ribosome-Associated Quality Control (RQC) Mechanism.Mol Neurobiol. 2021 Dec;58(12):6593-6609. doi: 10.1007/s12035-021-02564-x. Epub 2021 Sep 29. Mol Neurobiol. 2021. PMID: 34590243 Review.
-
HflX is a ribosome-splitting factor rescuing stalled ribosomes under stress conditions.Nat Struct Mol Biol. 2015 Nov;22(11):906-13. doi: 10.1038/nsmb.3103. Epub 2015 Oct 12. Nat Struct Mol Biol. 2015. PMID: 26458047
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources