

Significant determinants of mouse pain behaviour
- PMID: 25101983
- PMCID: PMC4125188
- DOI: 10.1371/journal.pone.0104458
Significant determinants of mouse pain behaviour
Erratum in
-
Correction: Significant Determinants of Mouse Pain Behaviour.PLoS One. 2021 Jan 15;16(1):e0245813. doi: 10.1371/journal.pone.0245813. eCollection 2021. PLoS One. 2021. PMID: 33449972 Free PMC article.
Abstract
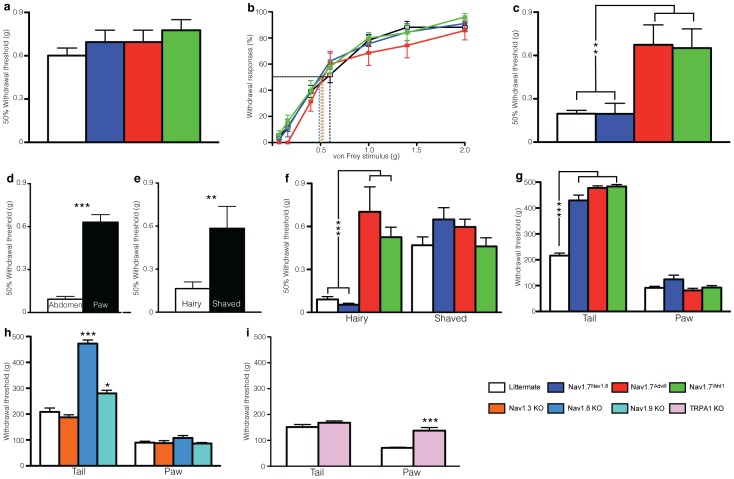
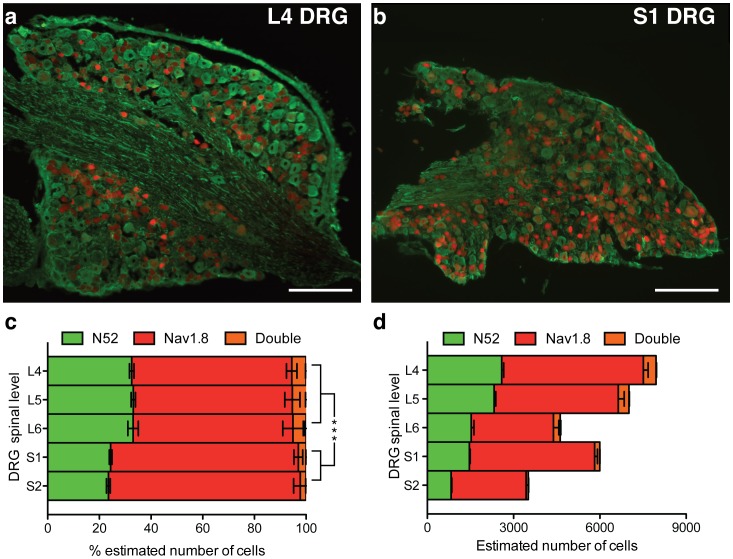
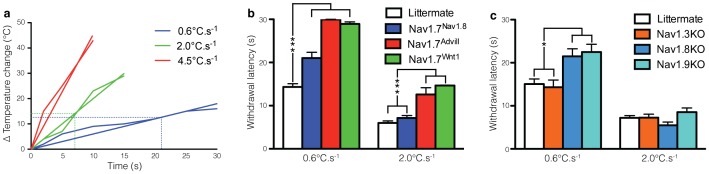
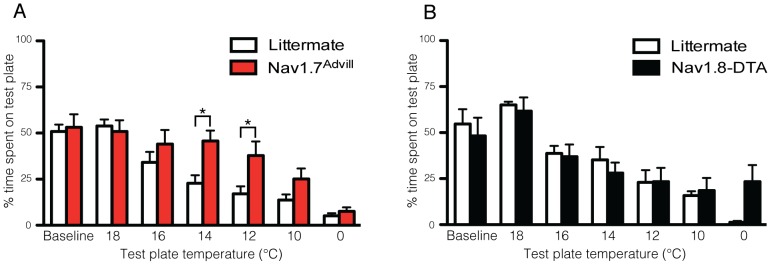
Transgenic mouse behavioural analysis has furthered our understanding of the molecular and cellular mechanisms underlying damage sensing and pain. However, it is not unusual for conflicting data on the pain phenotypes of knockout mice to be generated by reputable groups. Here we focus on some technical aspects of measuring mouse pain behaviour that are often overlooked, which may help explain discrepancies in the pain literature. We examined touch perception using von Frey hairs and mechanical pain thresholds using the Randall-Selitto test. Thermal pain thresholds were measured using the Hargreaves apparatus and a thermal place preference test. Sodium channel Nav1.7 knockout mice show a mechanical deficit in the hairy skin, but not the paw, whilst shaving the abdominal hair abolished this phenotype. Nav1.7, Nav1.8 and Nav1.9 knockout mice show deficits in noxious mechanosensation in the tail, but not the paw. TRPA1 knockout mice, however, have a loss of noxious mechanosensation in the paw but not the tail. Studies of heat and cold sensitivity also show variability depending on the intensity of the stimulus. Deleting Nav1.7, Nav1.8 or Nav1.9 in Nav1.8-positive sensory neurons attenuates responses to slow noxious heat ramps, whilst responses to fast noxious heat ramps are only reduced when Nav1.7 is lost in large diameter sensory neurons. Deleting Nav1.7 from all sensory neurons attenuates responses to noxious cooling but not extreme cold. Finally, circadian rhythms dramatically influence behavioural outcome measures such as von Frey responses, which change by 80% over the day. These observations demonstrate that fully characterising the phenotype of a transgenic mouse strain requires a range of behavioural pain models. Failure to conduct behavioural tests at different anatomical locations, stimulus intensities, and at different points in the circadian cycle may lead to a pain behavioural phenotype being misinterpreted, or missed altogether.
Conflict of interest statement
Figures

References
-
- Hargreaves K, Dubner R, Brown F, Flores C, Joris J (1988) A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 32: 77–88. - PubMed
-
- Randall LO, Selitto JJ (1957) A method for measurement of analgesic activity on inflamed tissue. Arch Int Pharmacodyn Ther 111: 409–419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
