

Syndecan-1 is required to maintain intradermal fat and prevent cold stress
- PMID: 25101993
- PMCID: PMC4125098
- DOI: 10.1371/journal.pgen.1004514
Syndecan-1 is required to maintain intradermal fat and prevent cold stress
Abstract
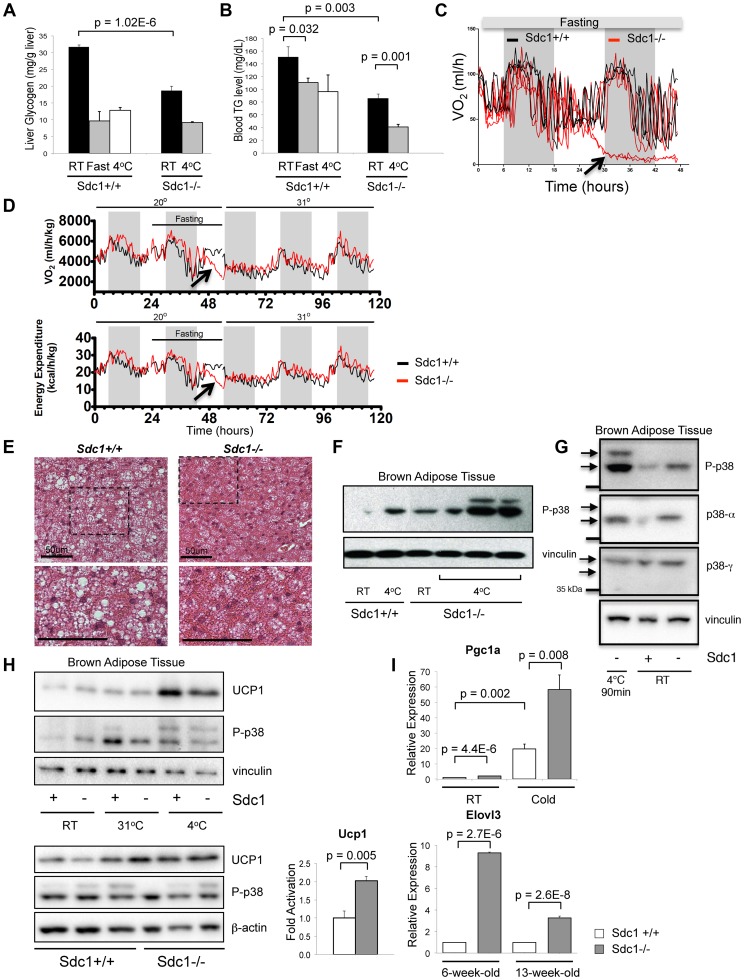
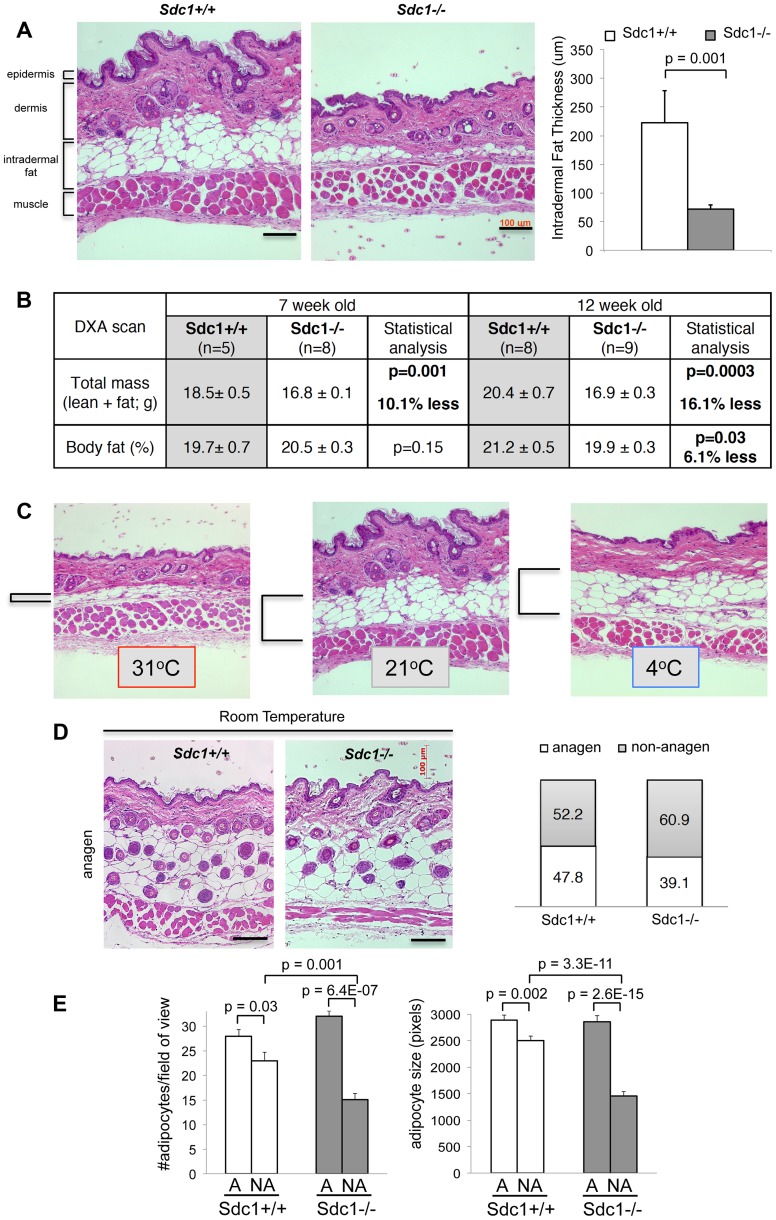
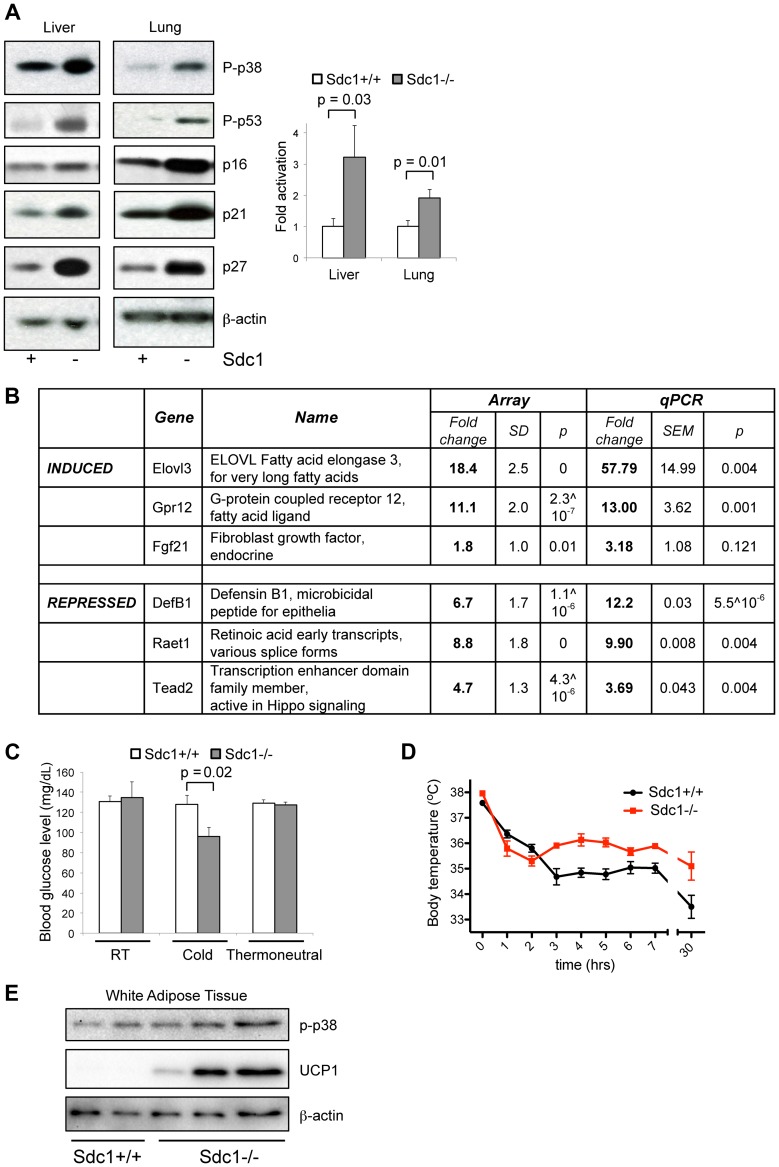
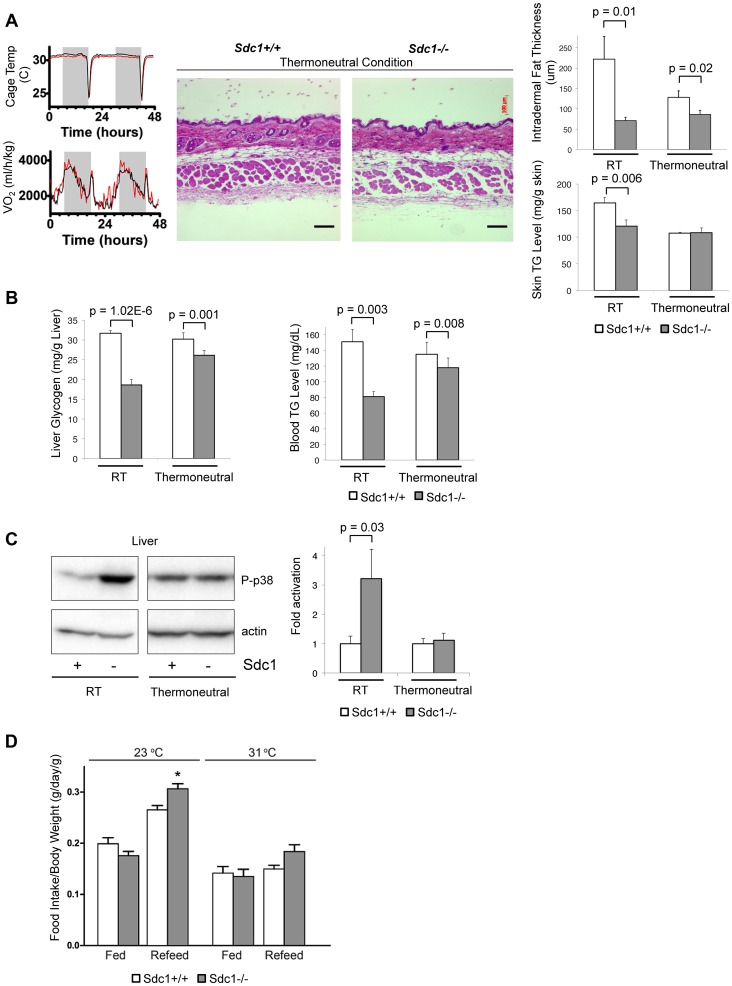
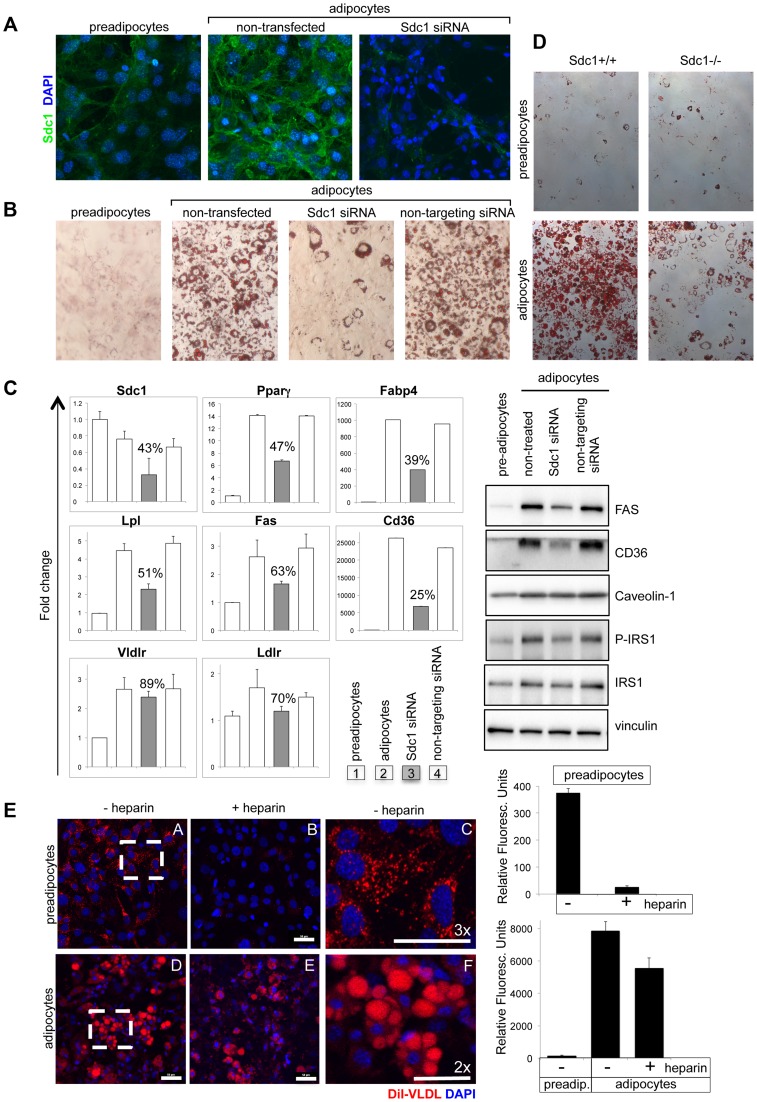
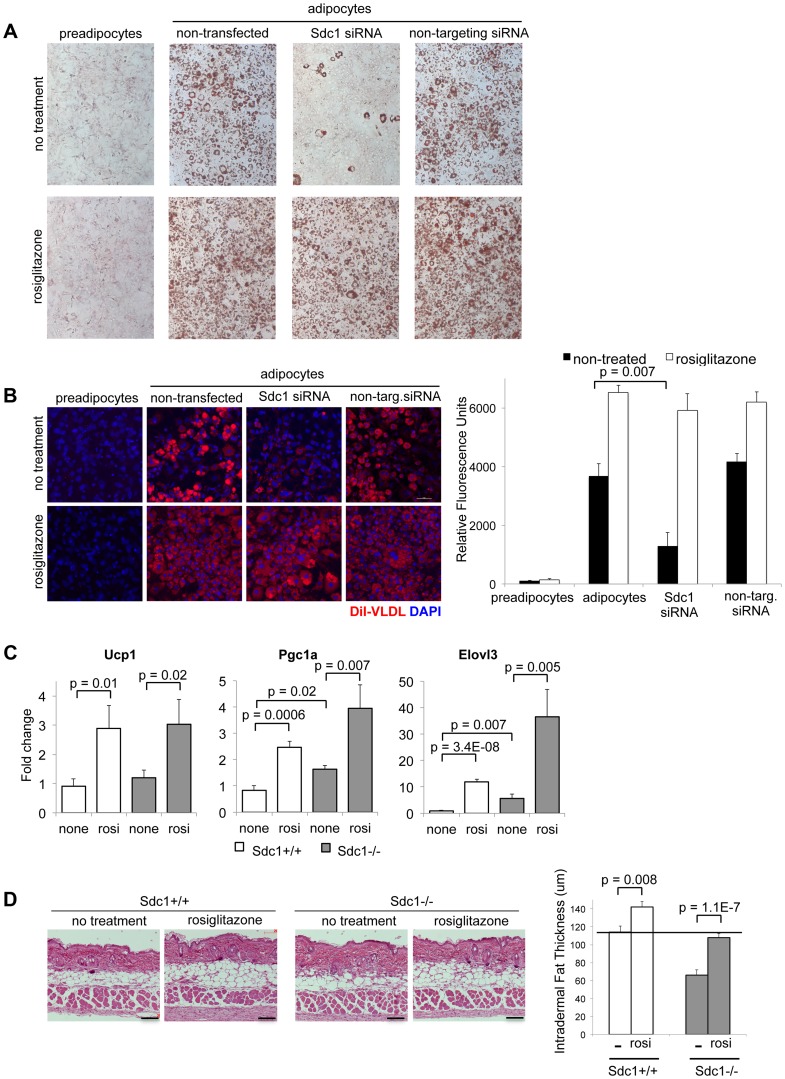
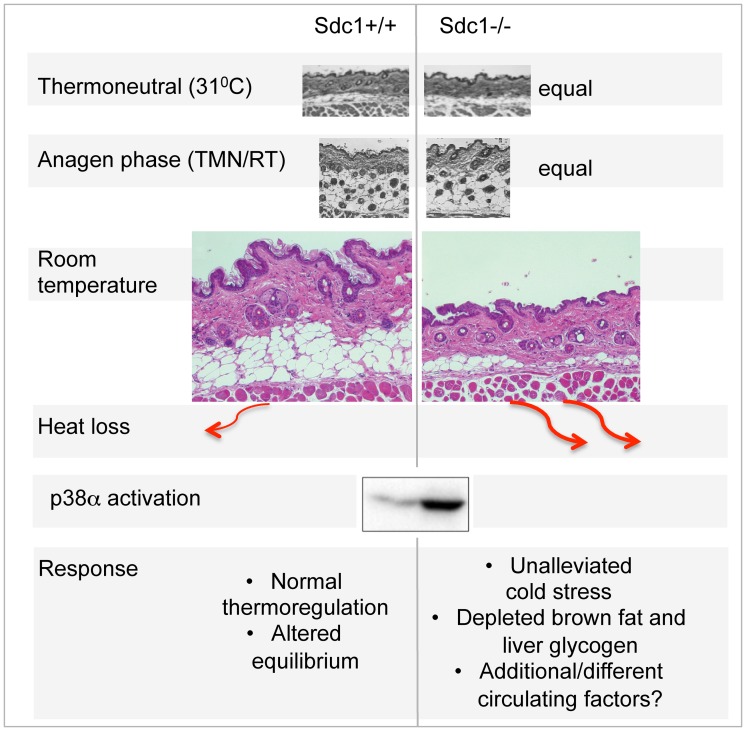
Homeostatic temperature regulation is fundamental to mammalian physiology and is controlled by acute and chronic responses of local, endocrine and nervous regulators. Here, we report that loss of the heparan sulfate proteoglycan, syndecan-1, causes a profoundly depleted intradermal fat layer, which provides crucial thermogenic insulation for mammals. Mice without syndecan-1 enter torpor upon fasting and show multiple indicators of cold stress, including activation of the stress checkpoint p38α in brown adipose tissue, liver and lung. The metabolic phenotype in mutant mice, including reduced liver glycogen, is rescued by housing at thermoneutrality, suggesting that reduced insulation in cool temperatures underlies the observed phenotypes. We find that syndecan-1, which functions as a facultative lipoprotein uptake receptor, is required for adipocyte differentiation in vitro. Intradermal fat shows highly dynamic differentiation, continuously expanding and involuting in response to hair cycle and ambient temperature. This physiology probably confers a unique role for Sdc1 in this adipocyte sub-type. The PPARγ agonist rosiglitazone rescues Sdc1-/- intradermal adipose tissue, placing PPARγ downstream of Sdc1 in triggering adipocyte differentiation. Our study indicates that disruption of intradermal adipose tissue development results in cold stress and complex metabolic pathology.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Golozoubova V, Gullberg H, Matthias A, Cannon B, Vennstrom B, et al. (2004) Depressed thermogenesis but competent brown adipose tissue recruitment in mice devoid of all hormone-binding thyroid hormone receptors. Molecular endocrinology 18: 384–401. - PubMed
-
- Overton JM, Williams TD (2004) Behavioral and physiologic responses to caloric restriction in mice. Physiology & behavior 81: 749–754. - PubMed
-
- Gordon CJ (2012) Thermal physiology of laboratory mice: Defining thermoneutrality. Journal of Thermal Biology 37: 654–685.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
