

The cohesin subunit Rad21 is required for synaptonemal complex maintenance, but not sister chromatid cohesion, during Drosophila female meiosis
- PMID: 25101996
- PMCID: PMC4125089
- DOI: 10.1371/journal.pgen.1004540
The cohesin subunit Rad21 is required for synaptonemal complex maintenance, but not sister chromatid cohesion, during Drosophila female meiosis
Abstract
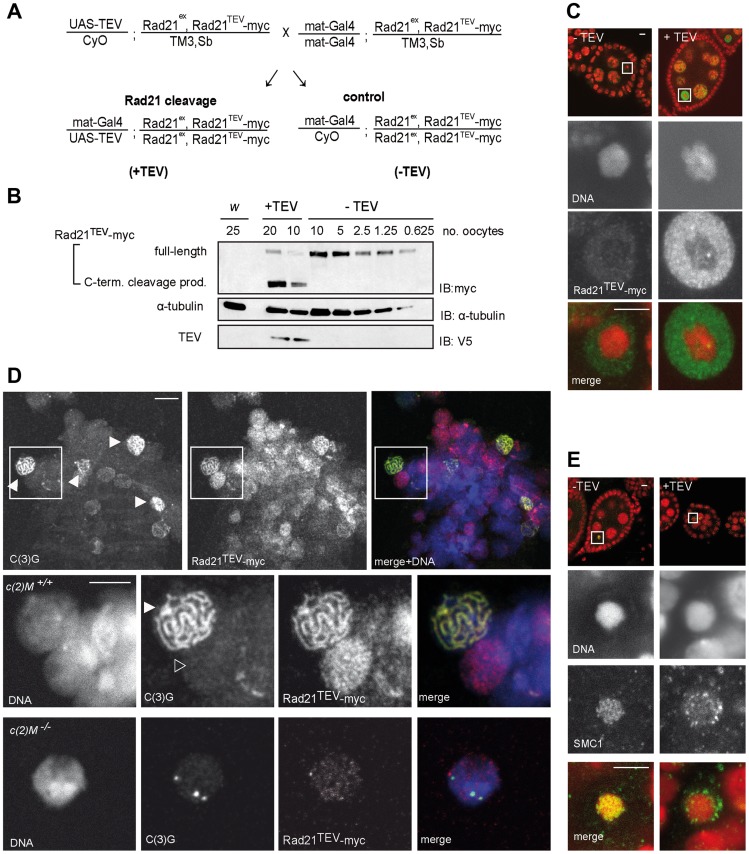
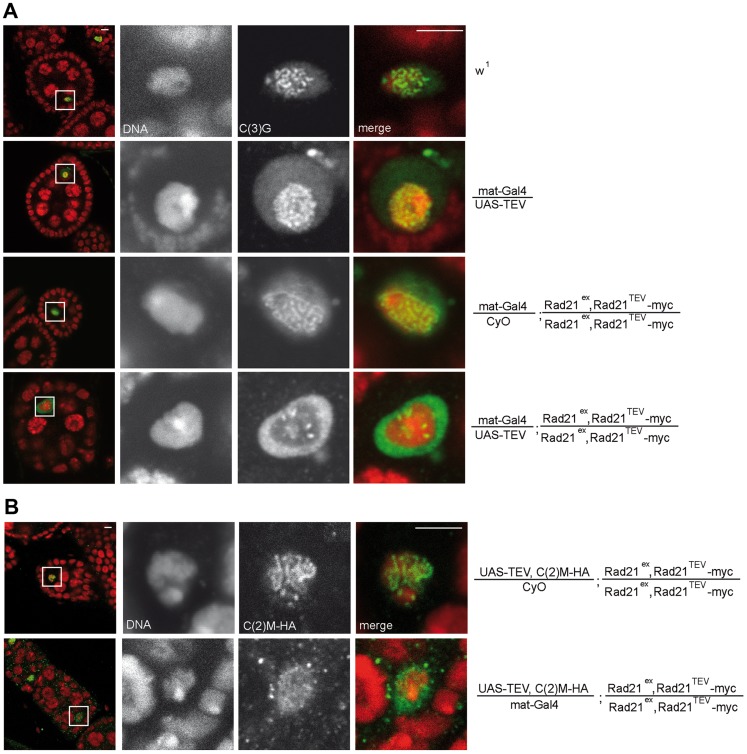
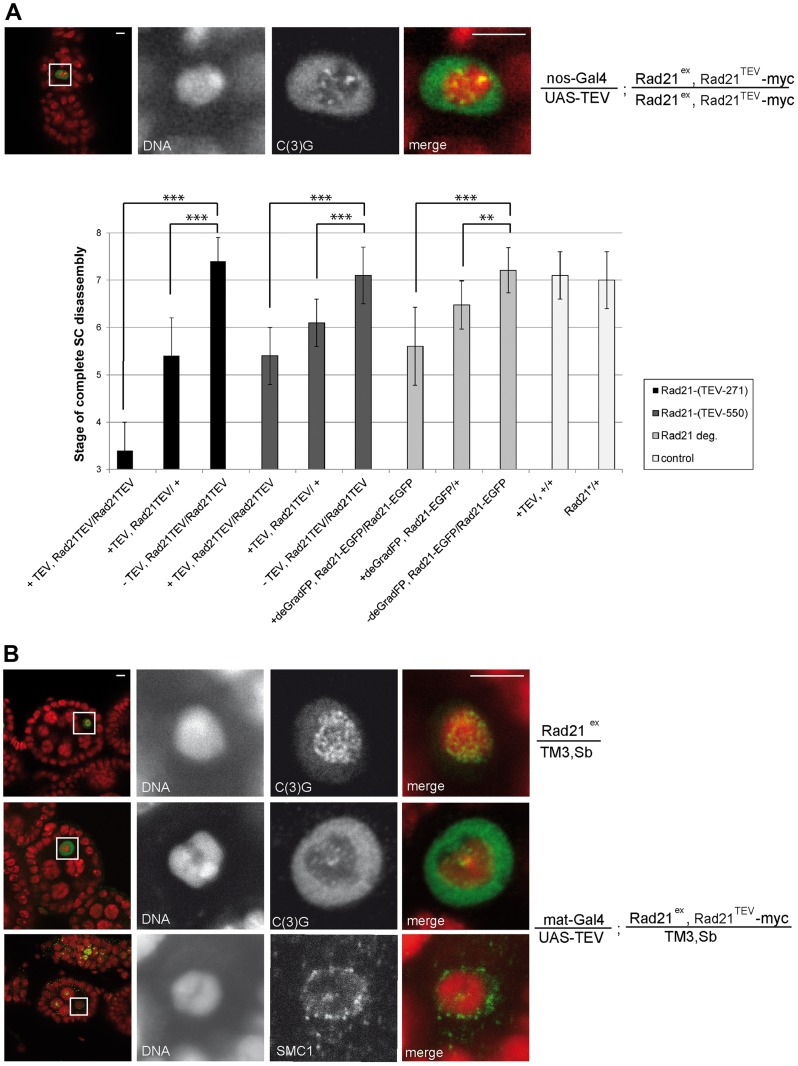
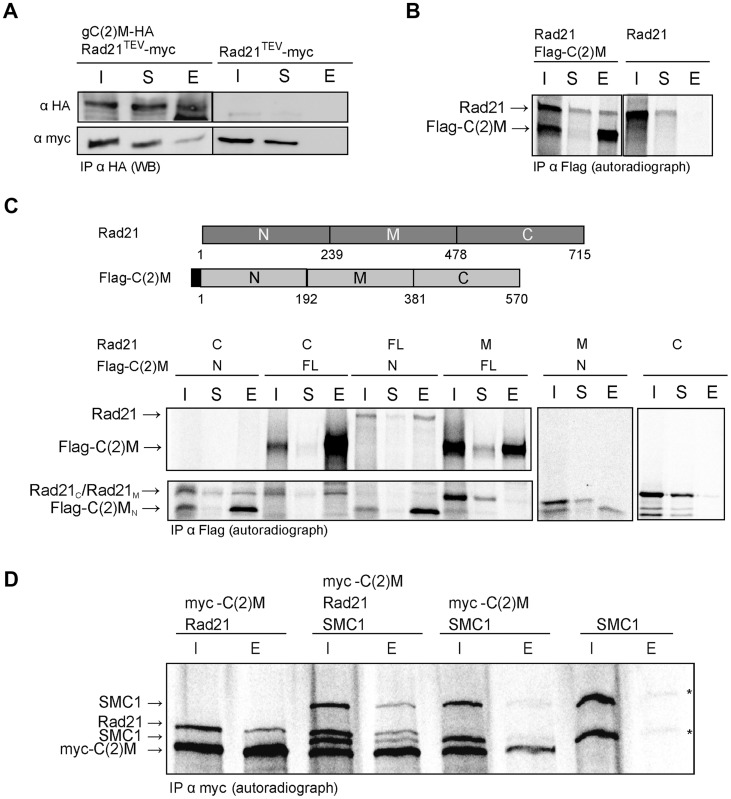
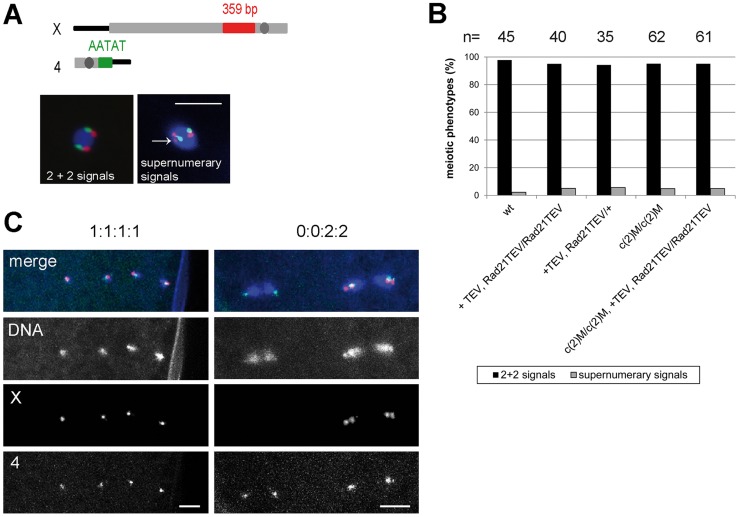
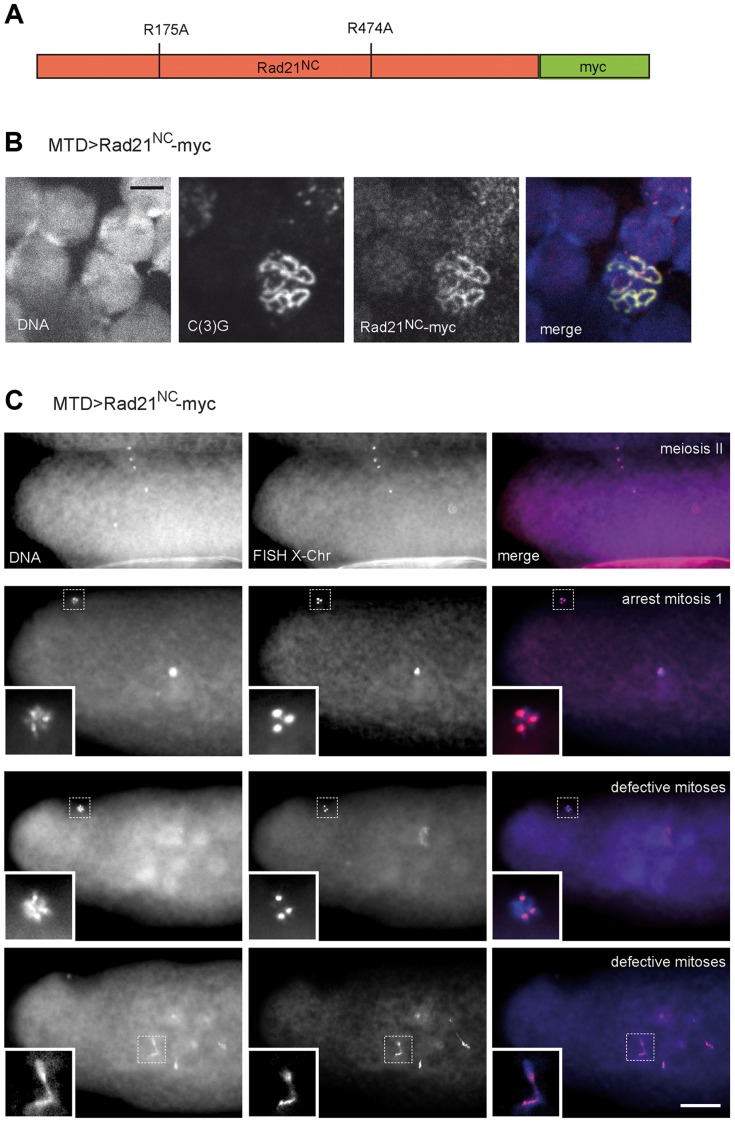
Replicated sister chromatids are held in close association from the time of their synthesis until their separation during the next mitosis. This association is mediated by the ring-shaped cohesin complex that appears to embrace the sister chromatids. Upon proteolytic cleavage of the α-kleisin cohesin subunit at the metaphase-to-anaphase transition by separase, sister chromatids are separated and segregated onto the daughter nuclei. The more complex segregation of chromosomes during meiosis is thought to depend on the replacement of the mitotic α-kleisin cohesin subunit Rad21/Scc1/Mcd1 by the meiotic paralog Rec8. In Drosophila, however, no clear Rec8 homolog has been identified so far. Therefore, we have analyzed the role of the mitotic Drosophila α-kleisin Rad21 during female meiosis. Inactivation of an engineered Rad21 variant by premature, ectopic cleavage during oogenesis results not only in loss of cohesin from meiotic chromatin, but also in precocious disassembly of the synaptonemal complex (SC). We demonstrate that the lateral SC component C(2)M can interact directly with Rad21, potentially explaining why Rad21 is required for SC maintenance. Intriguingly, the experimentally induced premature Rad21 elimination, as well as the expression of a Rad21 variant with destroyed separase consensus cleavage sites, do not interfere with chromosome segregation during meiosis, while successful mitotic divisions are completely prevented. Thus, chromatid cohesion during female meiosis does not depend on Rad21-containing cohesin.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The cohesion protein SOLO associates with SMC1 and is required for synapsis, recombination, homolog bias and cohesion and pairing of centromeres in Drosophila Meiosis.PLoS Genet. 2013;9(7):e1003637. doi: 10.1371/journal.pgen.1003637. Epub 2013 Jul 18. PLoS Genet. 2013. PMID: 23874232 Free PMC article.
-
Temporally and spatially selective loss of Rec8 protein from meiotic chromosomes during mammalian meiosis.J Cell Sci. 2003 Jul 1;116(Pt 13):2781-90. doi: 10.1242/jcs.00495. Epub 2003 May 20. J Cell Sci. 2003. PMID: 12759374
-
Separase Is Required for Homolog and Sister Disjunction during Drosophila melanogaster Male Meiosis, but Not for Biorientation of Sister Centromeres.PLoS Genet. 2016 Apr 27;12(4):e1005996. doi: 10.1371/journal.pgen.1005996. eCollection 2016 Apr. PLoS Genet. 2016. PMID: 27120695 Free PMC article.
-
Meiotic prophase-like pathway for cleavage-independent removal of cohesin for chromosome morphogenesis.Curr Genet. 2019 Aug;65(4):817-827. doi: 10.1007/s00294-019-00959-x. Epub 2019 Mar 28. Curr Genet. 2019. PMID: 30923890 Review.
-
The expanding phenotypes of cohesinopathies: one ring to rule them all!Cell Cycle. 2019 Nov;18(21):2828-2848. doi: 10.1080/15384101.2019.1658476. Epub 2019 Sep 13. Cell Cycle. 2019. PMID: 31516082 Free PMC article. Review.
Cited by
-
Female Meiosis: Synapsis, Recombination, and Segregation in Drosophila melanogaster.Genetics. 2018 Mar;208(3):875-908. doi: 10.1534/genetics.117.300081. Genetics. 2018. PMID: 29487146 Free PMC article. Review.
-
A cell cycle-independent, conditional gene inactivation strategy for differentially tagging wild-type and mutant cells.Elife. 2017 May 31;6:e26420. doi: 10.7554/eLife.26420. Elife. 2017. PMID: 28561736 Free PMC article.
-
De novo transcriptome assembly from the gonads of a scleractinian coral, Euphyllia ancora: molecular mechanisms underlying scleractinian gametogenesis.BMC Genomics. 2020 Oct 21;21(1):732. doi: 10.1186/s12864-020-07113-9. BMC Genomics. 2020. PMID: 33087060 Free PMC article.
-
Condensin I is required for faithful meiosis in Drosophila males.Chromosoma. 2020 Jun;129(2):141-160. doi: 10.1007/s00412-020-00733-w. Epub 2020 Apr 8. Chromosoma. 2020. PMID: 32314039 Free PMC article.
-
Regulated protein depletion by the auxin-inducible degradation system in Drosophila melanogaster.Fly (Austin). 2016 Jan 2;10(1):35-46. doi: 10.1080/19336934.2016.1168552. Epub 2016 Mar 24. Fly (Austin). 2016. PMID: 27010248 Free PMC article.
References
-
- Petronczki M, Siomos MF, Nasmyth K (2003) Un menage a quatre: the molecular biology of chromosome segregation in meiosis. Cell 112: 423–440. - PubMed
-
- McKim KS, Green-Marroquin BL, Sekelsky JJ, Chin G, Steinberg C, et al. (1998) Meiotic synapsis in the absence of recombination. Science 279: 876–878. - PubMed
-
- Manheim EA, McKim KS (2003) The Synaptonemal complex component C(2)M regulates meiotic crossing over in Drosophila. Curr Biol 13: 276–285. - PubMed
-
- Oliveira RA, Nasmyth K (2010) Getting through anaphase: splitting the sisters and beyond. Biochem Soc Trans 38: 1639–1644. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
