

C9orf72 repeat expansions cause neurodegeneration in Drosophila through arginine-rich proteins
- PMID: 25103406
- PMCID: PMC4944841
- DOI: 10.1126/science.1256800
C9orf72 repeat expansions cause neurodegeneration in Drosophila through arginine-rich proteins
Abstract
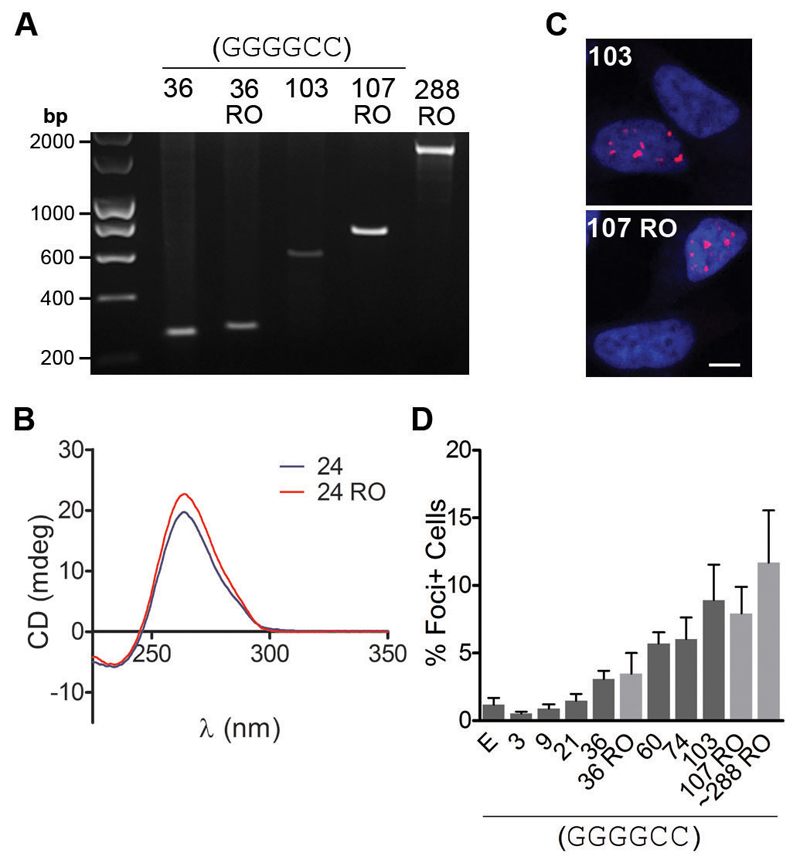
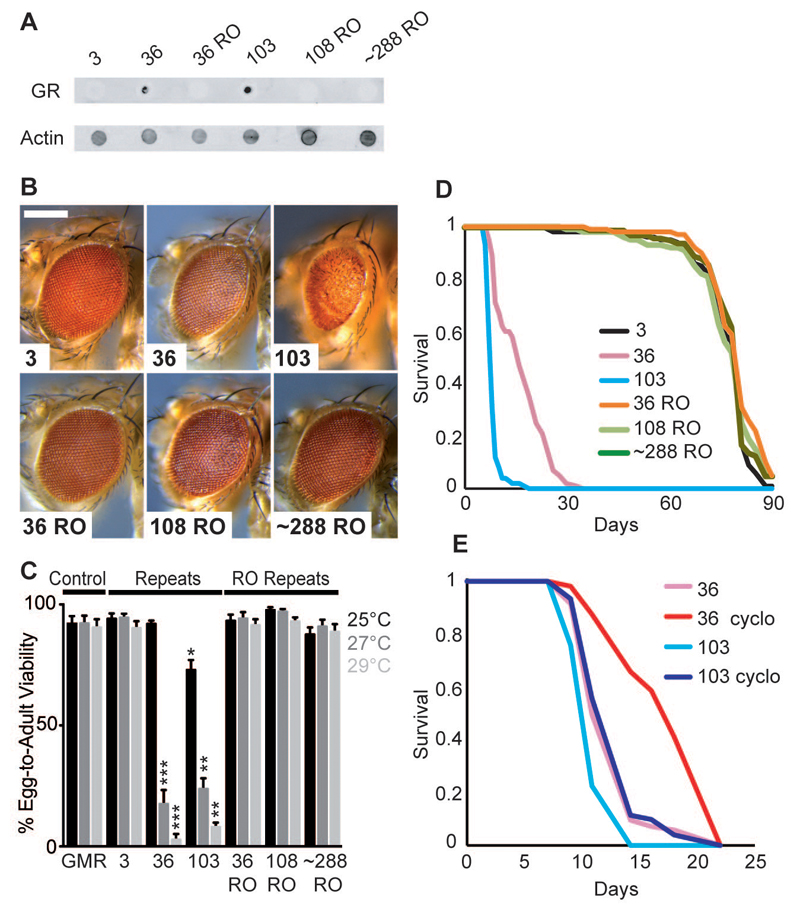
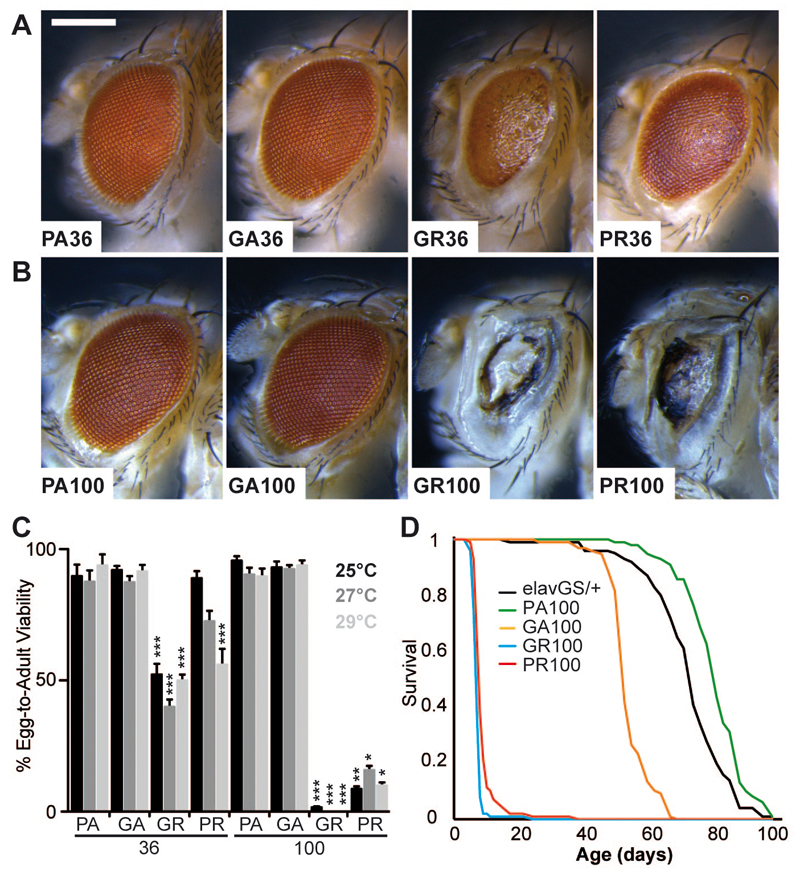
An expanded GGGGCC repeat in C9orf72 is the most common genetic cause of frontotemporal dementia and amyotrophic lateral sclerosis. A fundamental question is whether toxicity is driven by the repeat RNA itself and/or by dipeptide repeat proteins generated by repeat-associated, non-ATG translation. To address this question, we developed in vitro and in vivo models to dissect repeat RNA and dipeptide repeat protein toxicity. Expression of pure repeats, but not stop codon-interrupted "RNA-only" repeats in Drosophila caused adult-onset neurodegeneration. Thus, expanded repeats promoted neurodegeneration through dipeptide repeat proteins. Expression of individual dipeptide repeat proteins with a non-GGGGCC RNA sequence revealed that both poly-(glycine-arginine) and poly-(proline-arginine) proteins caused neurodegeneration. These findings are consistent with a dual toxicity mechanism, whereby both arginine-rich proteins and repeat RNA contribute to C9orf72-mediated neurodegeneration.
Copyright © 2014, American Association for the Advancement of Science.
Figures
Comment in
-
Cell Biology. Clogging information flow in ALS.Science. 2014 Sep 5;345(6201):1118-9. doi: 10.1126/science.1259461. Science. 2014. PMID: 25190778 No abstract available.
References
-
- Lillo P, Hodges JR. Frontotemporal dementia and motor neurone disease: overlapping clinic-pathological disorders. J Clin Neurosci. 2009;16:1131–1135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
