

Tyr-301 phosphorylation inhibits pyruvate dehydrogenase by blocking substrate binding and promotes the Warburg effect
- PMID: 25104357
- PMCID: PMC4176253
- DOI: 10.1074/jbc.M114.593970
Tyr-301 phosphorylation inhibits pyruvate dehydrogenase by blocking substrate binding and promotes the Warburg effect
Abstract
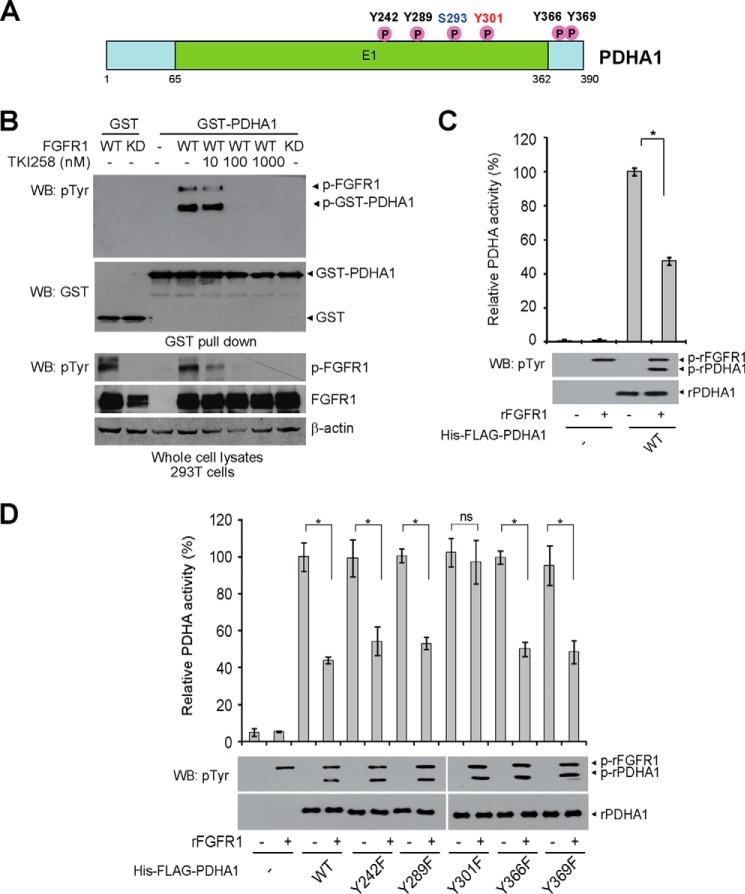
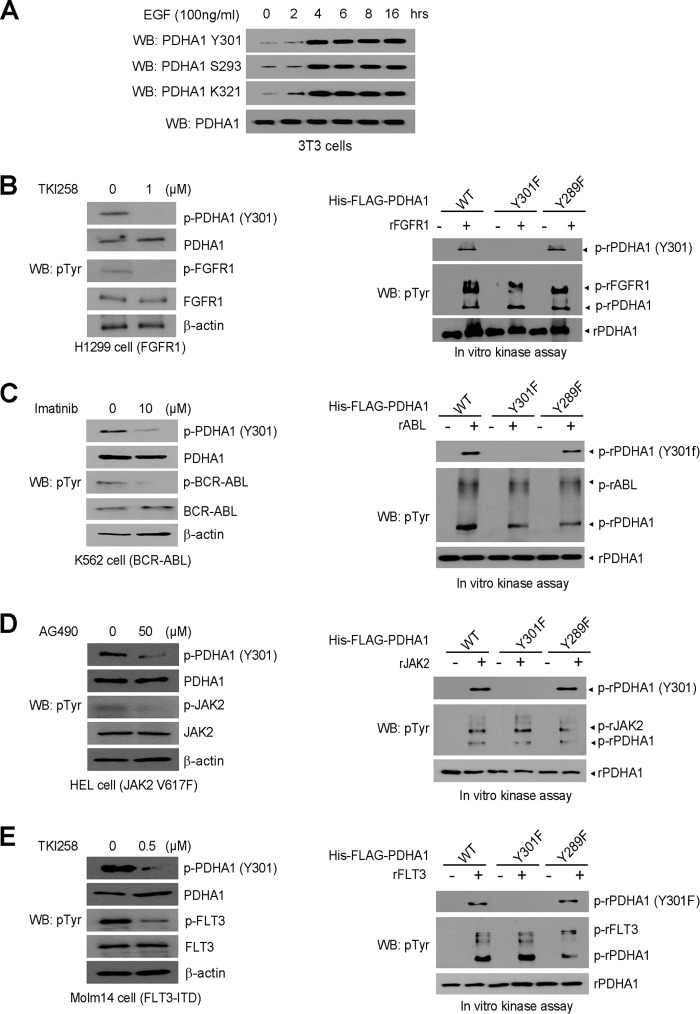
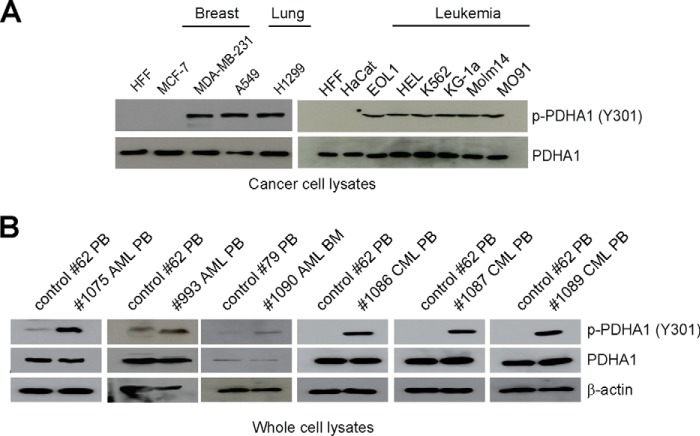
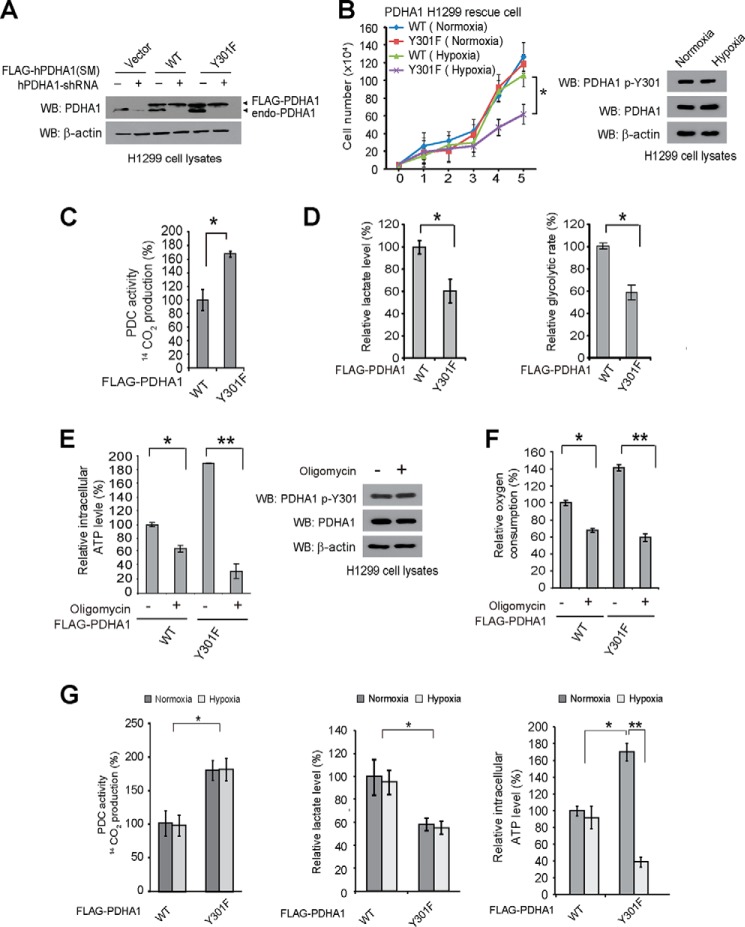
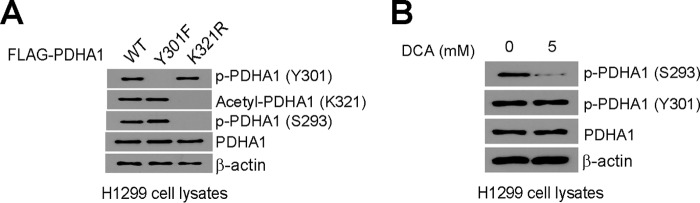
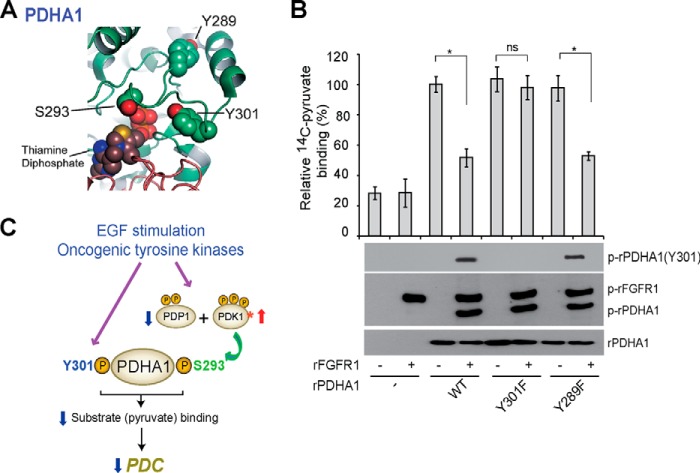
The mitochondrial pyruvate dehydrogenase complex (PDC) plays a crucial role in regulation of glucose homoeostasis in mammalian cells. PDC flux depends on catalytic activity of the most important enzyme component pyruvate dehydrogenase (PDH). PDH kinase inactivates PDC by phosphorylating PDH at specific serine residues, including Ser-293, whereas dephosphorylation of PDH by PDH phosphatase restores PDC activity. The current understanding suggests that Ser-293 phosphorylation of PDH impedes active site accessibility to its substrate pyruvate. Here, we report that phosphorylation of a tyrosine residue Tyr-301 also inhibits PDH α 1 (PDHA1) by blocking pyruvate binding through a novel mechanism in addition to Ser-293 phosphorylation. In addition, we found that multiple oncogenic tyrosine kinases directly phosphorylate PDHA1 at Tyr-301, and Tyr-301 phosphorylation of PDHA1 is common in EGF-stimulated cells as well as diverse human cancer cells and primary leukemia cells from human patients. Moreover, expression of a phosphorylation-deficient PDHA1 Y301F mutant in cancer cells resulted in increased oxidative phosphorylation, decreased cell proliferation under hypoxia, and reduced tumor growth in mice. Together, our findings suggest that phosphorylation at distinct serine and tyrosine residues inhibits PDHA1 through distinct mechanisms to impact active site accessibility, which act in concert to regulate PDC activity and promote the Warburg effect.
Keywords: Cell Proliferation; Phosphotyrosine Signaling; Pyruvate Dehydrogenase Complex (PDC); Tumor Metabolism; Warburg Effect.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Cairns R. A., Harris I. S., Mak T. W. (2011) Regulation of cancer cell metabolism. Nat. Rev. Cancer 11, 85–95 - PubMed
-
- Kroemer G., Pouyssegur J. (2008) Tumor cell metabolism: cancer's Achilles' heel. Cancer Cell 13, 472–482 - PubMed
-
- Hiromasa Y., Fujisawa T., Aso Y., Roche T. E. (2004) Organization of the cores of the mammalian pyruvate dehydrogenase complex formed by E2 and E2 plus the E3-binding protein and their capacities to bind the E1 and E3 components. J. Biol. Chem. 279, 6921–6933 - PubMed
-
- Read R. J. (2001) Pushing the boundaries of molecular replacement with maximum likelihood. Acta Crystallogr. D 57, 1373–1382 - PubMed
-
- Harris R. A., Bowker-Kinley M. M., Huang B., Wu P. (2002) Regulation of the activity of the pyruvate dehydrogenase complex. Adv. Enzyme Regul. 42, 249–259 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
