

Molecular phylogeny and intricate evolutionary history of the three isofunctional enzymes involved in the oxidation of protoporphyrinogen IX
- PMID: 25108393
- PMCID: PMC4231631
- DOI: 10.1093/gbe/evu170
Molecular phylogeny and intricate evolutionary history of the three isofunctional enzymes involved in the oxidation of protoporphyrinogen IX
Abstract
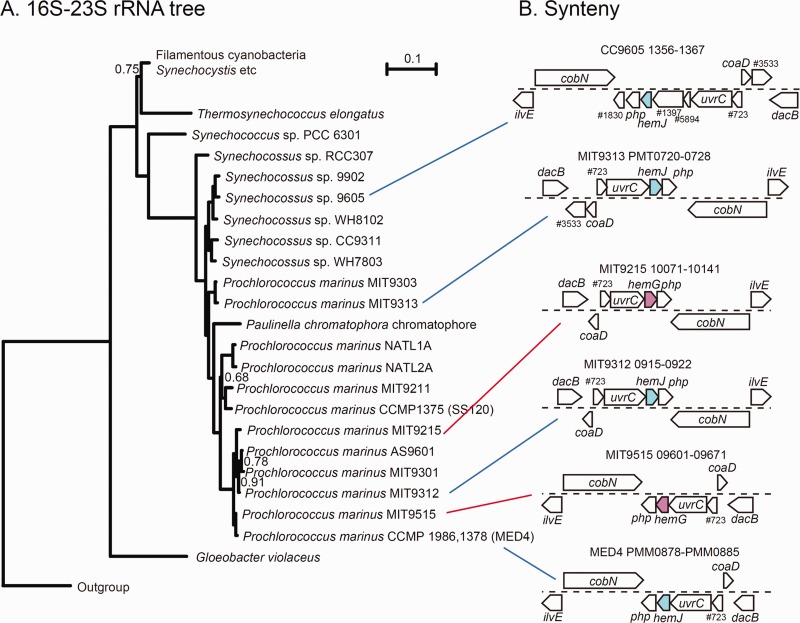
Tetrapyrroles such as heme and chlorophyll are essential for biological processes, including oxygenation, respiration, and photosynthesis. In the tetrapyrrole biosynthesis pathway, protoporphyrinogen IX oxidase (Protox) catalyzes the formation of protoporphyrin IX, the last common intermediate for the biosynthesis of heme and chlorophyll. Three nonhomologous isofunctional enzymes, HemG, HemJ, and HemY, for Protox have been identified. To reveal the distribution and evolution of the three Protox enzymes, we identified homologs of each along with other heme biosynthetic enzymes by whole-genome clustering across three domains of life. Most organisms possess only one of the three Protox types, with some exceptions. Detailed phylogenetic analysis revealed that HemG is mostly limited to γ-Proteobacteria whereas HemJ may have originated within α-Proteobacteria and transferred to other Proteobacteria and Cyanobacteria. In contrast, HemY is ubiquitous in prokaryotes and is the only Protox in eukaryotes, so this type may be the ancestral Protox. Land plants have a unique HemY homolog that is also shared by Chloroflexus species, in addition to the main HemY homolog originating from Cyanobacteria. Meanwhile, organisms missing any Protox can be classified into two groups; those lacking most heme synthetic genes, which necessarily depend on external heme supply, and those lacking only genes involved in the conversion of uroporphyrinogen III into heme, which would use a precorrin2-dependent alternative pathway. However, hemN encoding coproporphyrinogen IX oxidase was frequently found in organisms lacking Protox enzyme, which suggests a unique role of this gene other than in heme biosynthesis.
Keywords: HemG; HemJ; HemY; heme; protoporphyrin IX; tetrapyrrole.
© The Author(s) 2014. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures





References
-
- Abicht HK, Martinez J, Layer G, Jahn D, Solioz M. Lactococcus lactis HemW (HemN) is a haem-binding protein with a putative role in haem trafficking. Biochem J. 2012;442:335–343. - PubMed
-
- Adachi J, Hasegawa M. 1996. MOLPHY Version 2.3: programs for molecular phylogenetics based on maximum likelihood. Computer Science Monographs. Vol. 28. Tokyo (Japan): Institute of Statistical Mathematics. p. 1–150.
-
- Camadro JM, Labbe P. Cloning and characterization of the yeast HEM14 gene coding for protoporphyrinogen oxidase, the molecular target of diphenyl ether-type herbicides. J Biol Chem. 1996;271:9120–9128. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
