

An acetate switch regulates stress erythropoiesis
- PMID: 25108527
- PMCID: PMC4159437
- DOI: 10.1038/nm.3587
An acetate switch regulates stress erythropoiesis
Abstract
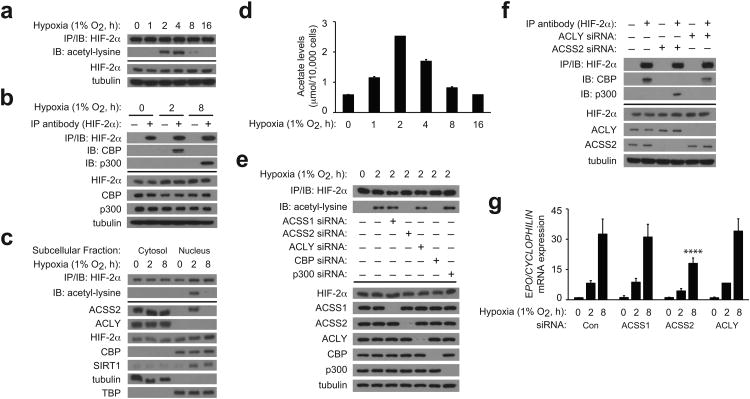
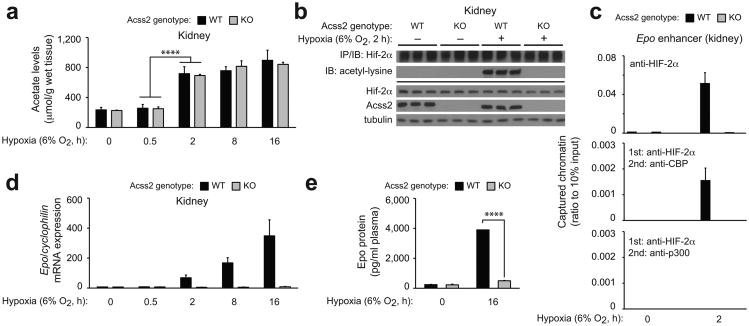
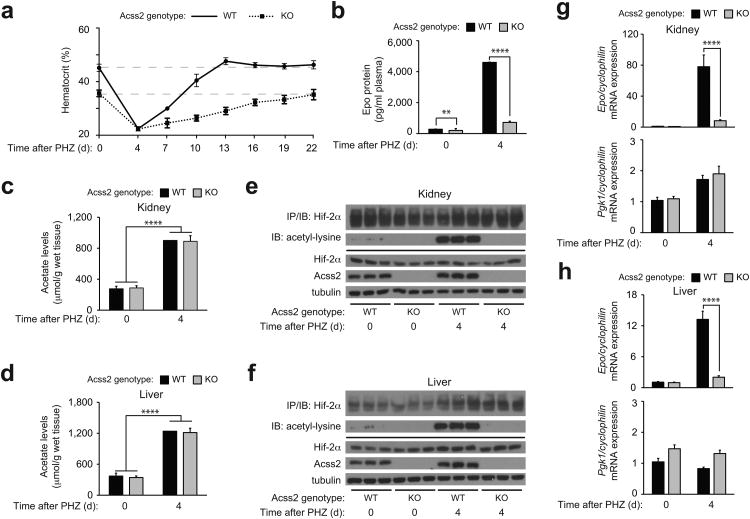
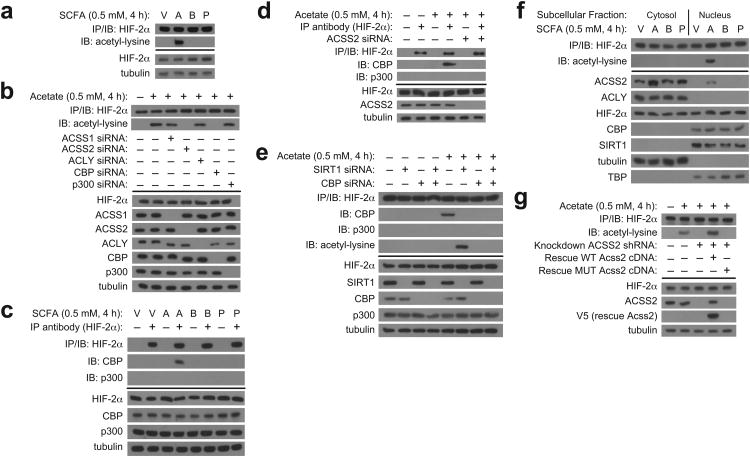
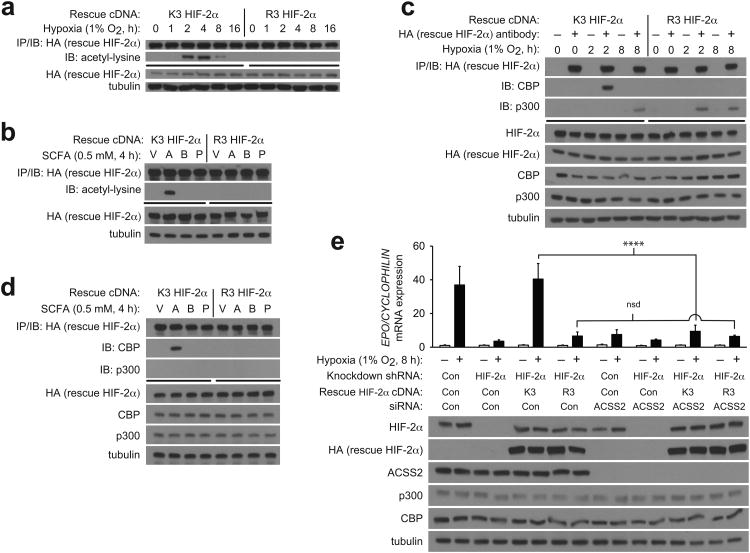
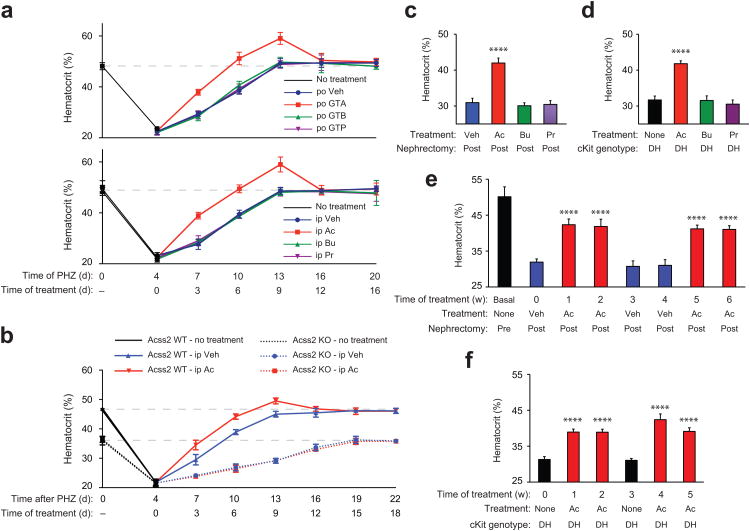
The hormone erythropoietin (EPO), which is synthesized in the kidney or liver of adult mammals, controls erythrocyte production and is regulated by the stress-responsive transcription factor hypoxia-inducible factor-2 (HIF-2). We previously reported that the lysine acetyltransferase CREB-binding protein (CBP) is required for HIF-2α acetylation and efficient HIF-2-dependent EPO induction during hypoxia. We now show that these processes require acetate-dependent acetyl CoA synthetase 2 (ACSS2). In human Hep3B hepatoma cells and in EPO-generating organs of hypoxic or acutely anemic mice, acetate levels rise and ACSS2 is required for HIF-2α acetylation, CBP-HIF-2α complex formation, CBP-HIF-2α recruitment to the EPO enhancer and efficient induction of EPO gene expression. In acutely anemic mice, acetate supplementation augments stress erythropoiesis in an ACSS2-dependent manner. Moreover, in acquired and inherited chronic anemia mouse models, acetate supplementation increases EPO expression and the resting hematocrit. Thus, a mammalian stress-responsive acetate switch controls HIF-2 signaling and EPO induction during pathophysiological states marked by tissue hypoxia.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Bunn HF, Poyton RO. Oxygen sensing and molecular adaptation to hypoxia. Physiological Reviews. 1996;76:839–885. - PubMed
-
- Richalet JP. Oxygen sensors in the organism: examples of regulation under altitude hypoxia in mammals. Comparative Biochemistry & Physiology, Part A Physiology. 1997;118:9–14. - PubMed
-
- Koury MJ. Erythropoietin: the story of hypoxia and a finely regulated hematopoietic hormone. Exp Hematol. 2005;33:1263–1270. - PubMed
-
- Semenza GL. Regulation of oxygen homeostasis by hypoxia-inducible factor 1. Physiology (Bethesda) 2009;24:97–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
