

Functional RNAi screen targeting cytokine and growth factor receptors reveals oncorequisite role for interleukin-2 gamma receptor in JAK3-mutation-positive leukemia
- PMID: 25109334
- PMCID: PMC4324389
- DOI: 10.1038/onc.2014.243
Functional RNAi screen targeting cytokine and growth factor receptors reveals oncorequisite role for interleukin-2 gamma receptor in JAK3-mutation-positive leukemia
Abstract
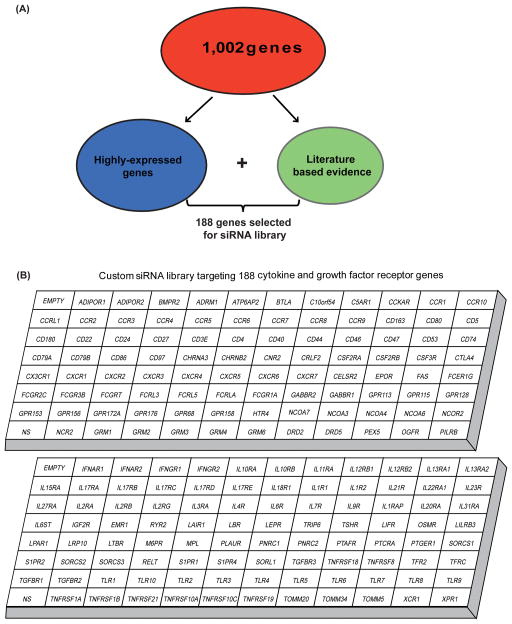
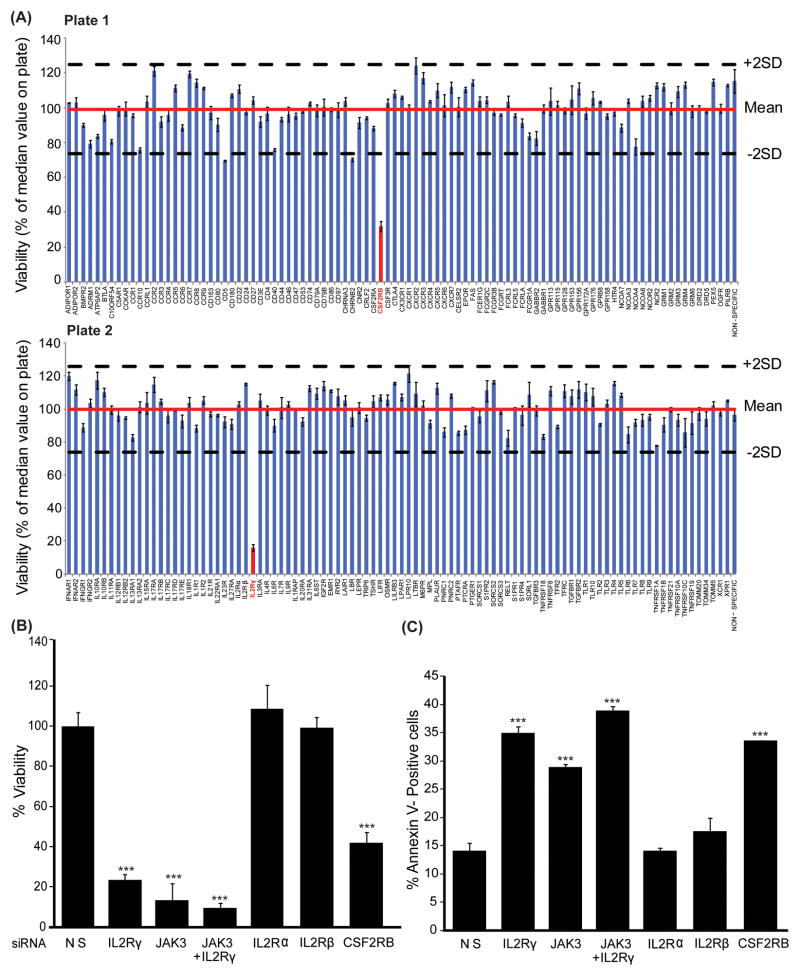
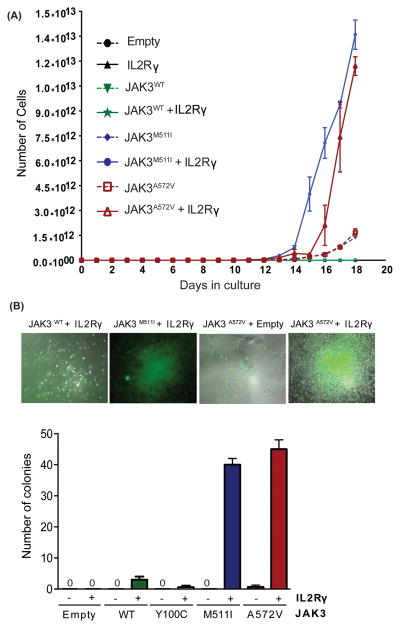
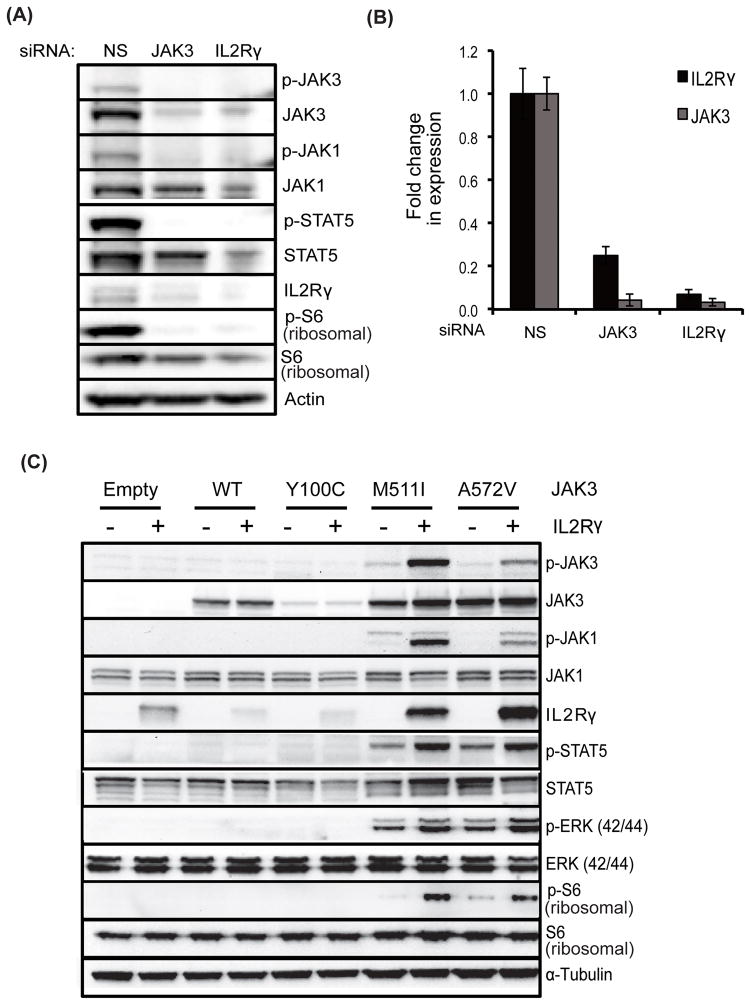
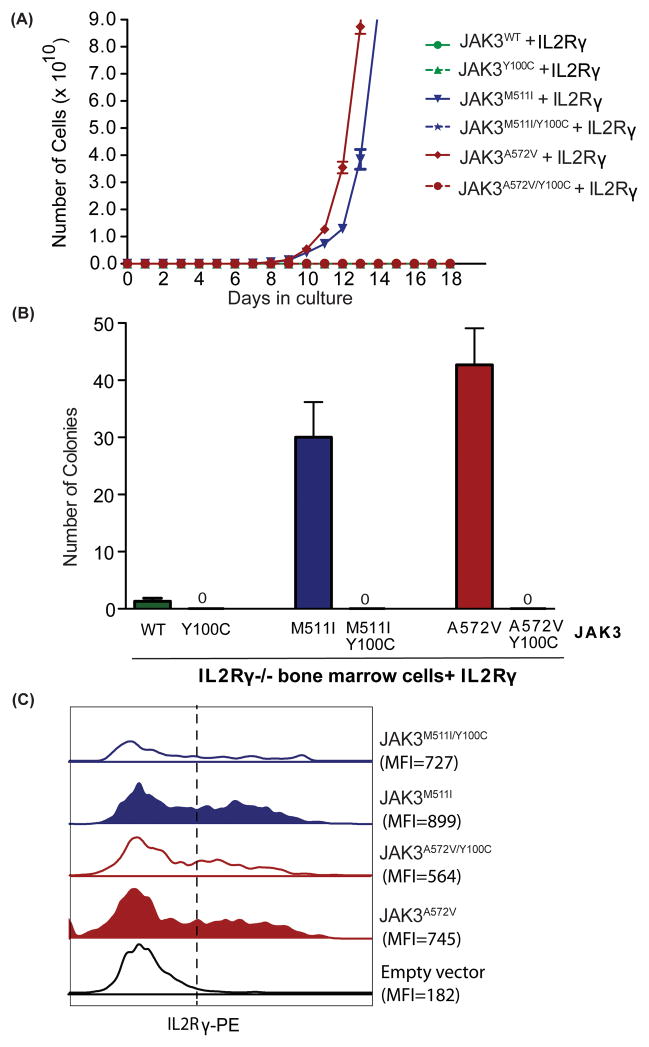
To understand the role of cytokine and growth factor receptor-mediated signaling in leukemia pathogenesis, we designed a functional RNA interference (RNAi) screen targeting 188 cytokine and growth factor receptors that we found highly expressed in primary leukemia specimens. Using this screen, we identified interleukin-2 gamma receptor (IL2Rγ) as a critical growth determinant for a JAK3(A572V) mutation-positive acute myeloid leukemia cell line. We observed that knockdown of IL2Rγ abrogates phosphorylation of JAK3 and downstream signaling molecules, JAK1, STAT5, MAPK and pS6 ribosomal protein. Overexpression of IL2Rγ in murine cells increased the transforming potential of activating JAK3 mutations, whereas absence of IL2Rγ completely abrogated the clonogenic potential of JAK3(A572V), as well as the transforming potential of additional JAK3-activating mutations such as JAK3(M511I). In addition, mutation at the IL2Rγ interaction site in the FERM domain of JAK3 (Y100C) completely abrogated JAK3-mediated leukemic transformation. Mechanistically, we found IL2Rγ contributes to constitutive JAK3 mutant signaling by increasing JAK3 expression and phosphorylation. Conversely, we found that mutant, but not wild-type JAK3, increased the expression of IL2Rγ, indicating IL2Rγ and JAK3 contribute to constitutive JAK/STAT signaling through their reciprocal regulation. Overall, we demonstrate a novel role for IL2Rγ in potentiating oncogenesis in the setting of JAK3-mutation-positive leukemia. In addition, our study highlights an RNAi-based functional assay that can be used to facilitate the identification of non-kinase cytokine and growth factor receptor targets for inhibiting leukemic cell growth.
Conflict of interest statement
OHSU and B.J.D. have a financial interest in Molecular MD (5% equity or less). This potential conflict of interest has been reviewed and managed by the OHSU Conflict of Interest in Research Committee and the Integrity Program Oversight Council. OHSU has clinical trial contracts with Novartis, and Bristol Myers Squibb to pay for patient costs, nurse and data manager salaries, and institutional overhead. B.J.D. does not derive salary, nor do their laboratories receive funds, from these contracts. B.J.D serves as a consultant for Roche and Nodality (Consulting income $10,000 or over).
Figures
References
-
- Estey E, Dohner H. Acute myeloid leukaemia. Lancet. 2006 Nov 25;368(9550):1894–907. eng. - PubMed
-
- Graf M, Hecht K, Reif S, Pelka-Fleischer R, Pfister K, Schmetzer H. Expression and prognostic value of hemopoietic cytokine receptors in acute myeloid leukemia (AML): implications for future therapeutical strategies. Eur J Haematol. 2004 Feb;72(2):89–106. eng. - PubMed
-
- Vainchenker W, Constantinescu SN. JAK/STAT signaling in hematological malignancies. Oncogene. 2013 May 23;32(21):2601–13. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
