

Structure and dynamics of AMPA receptor GluA2 in resting, pre-open, and desensitized states
- PMID: 25109876
- PMCID: PMC4263325
- DOI: 10.1016/j.cell.2014.07.023
Structure and dynamics of AMPA receptor GluA2 in resting, pre-open, and desensitized states
Abstract
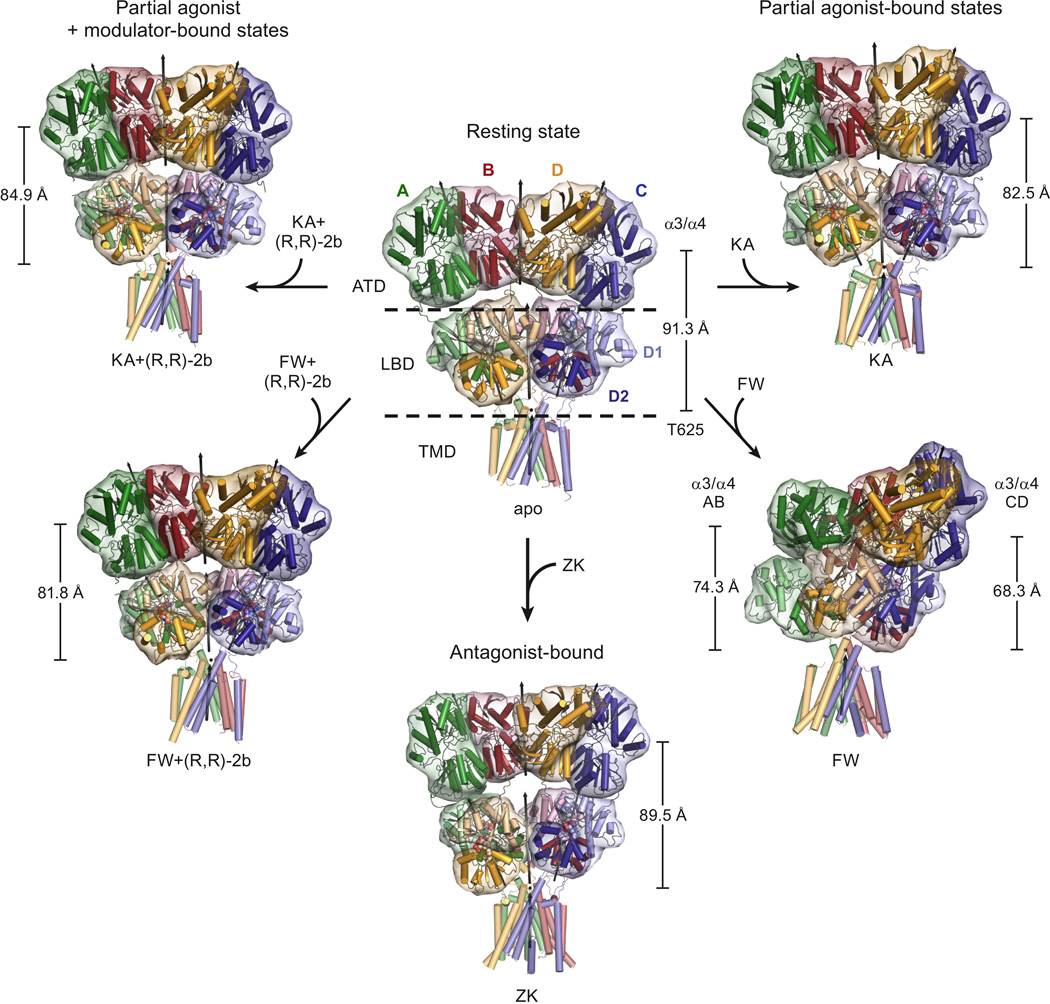
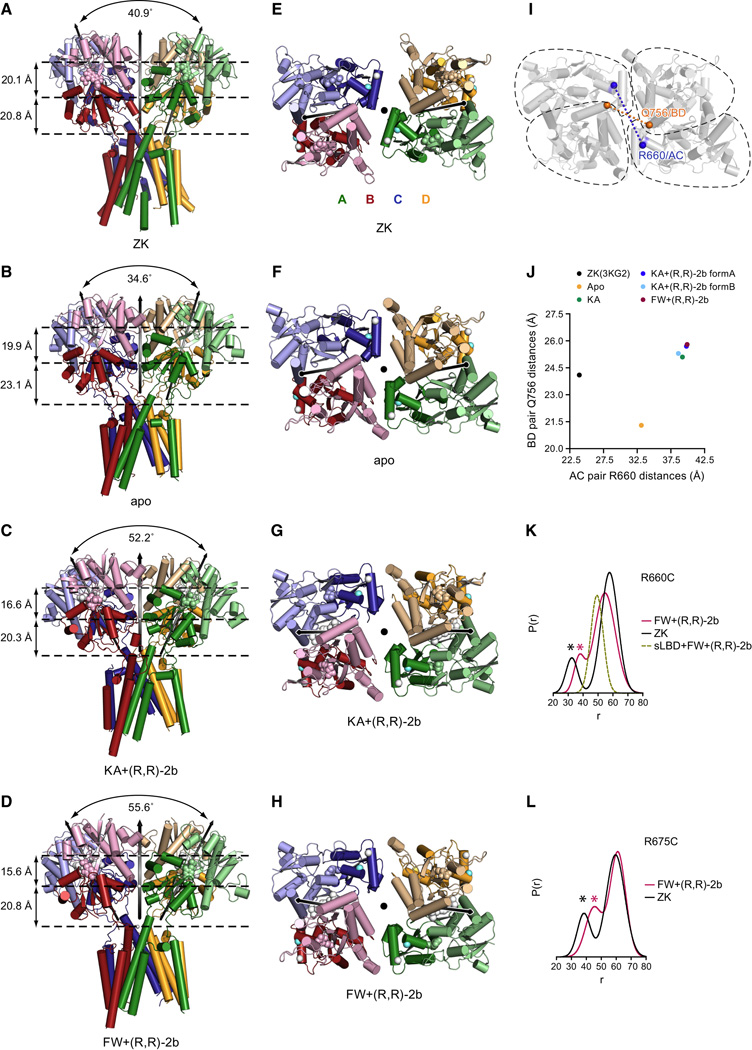
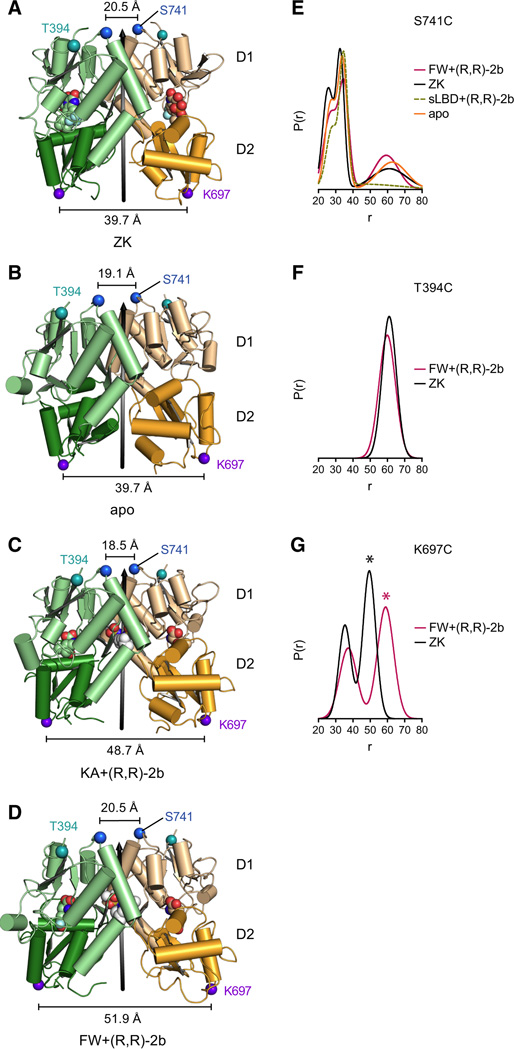
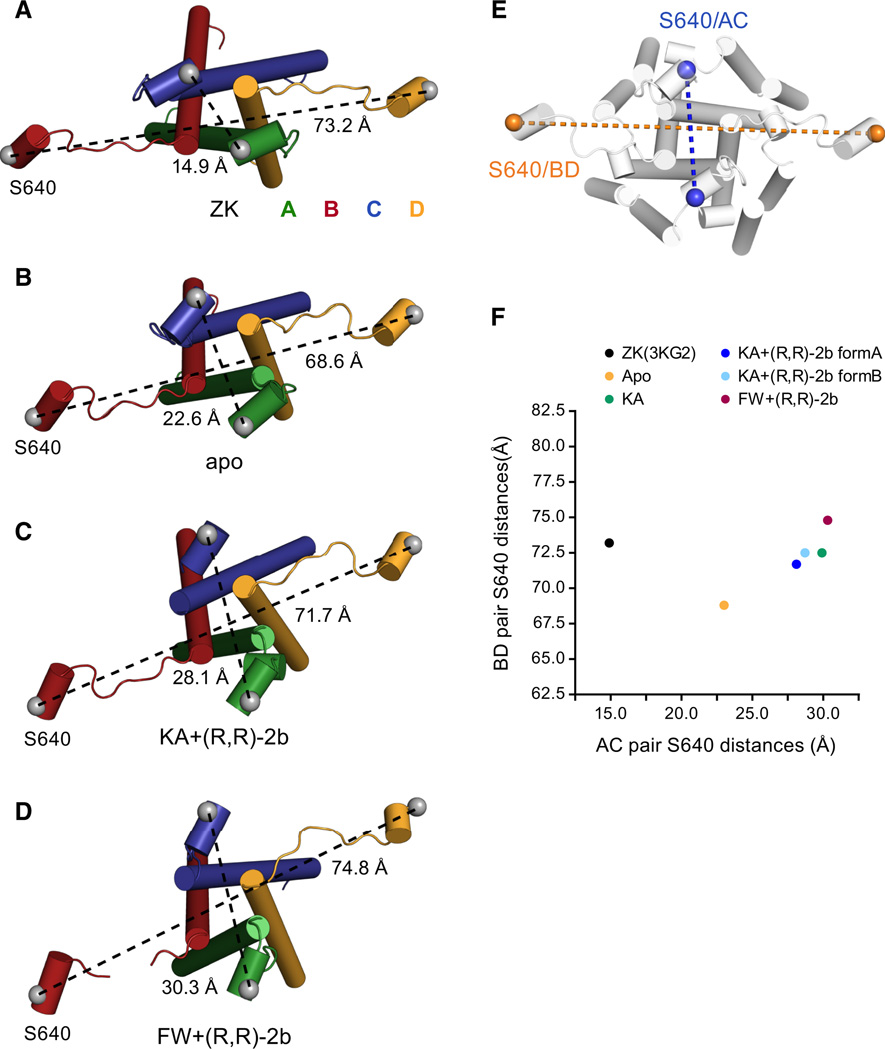
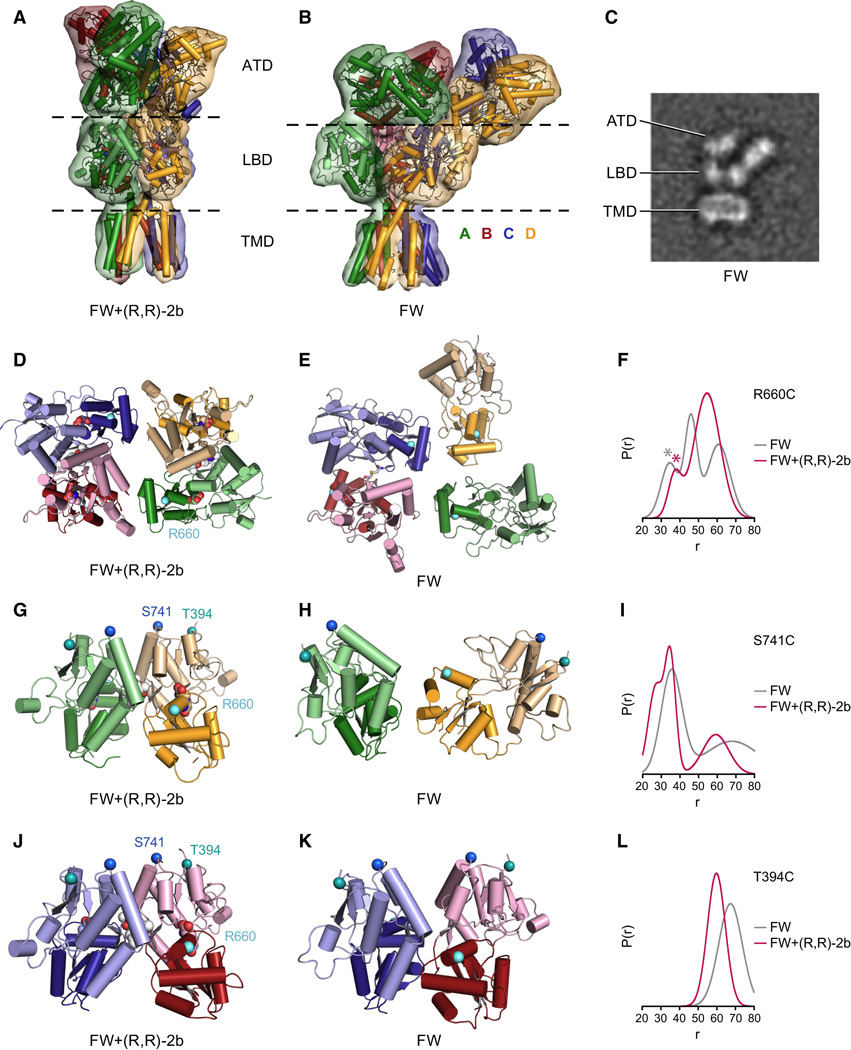
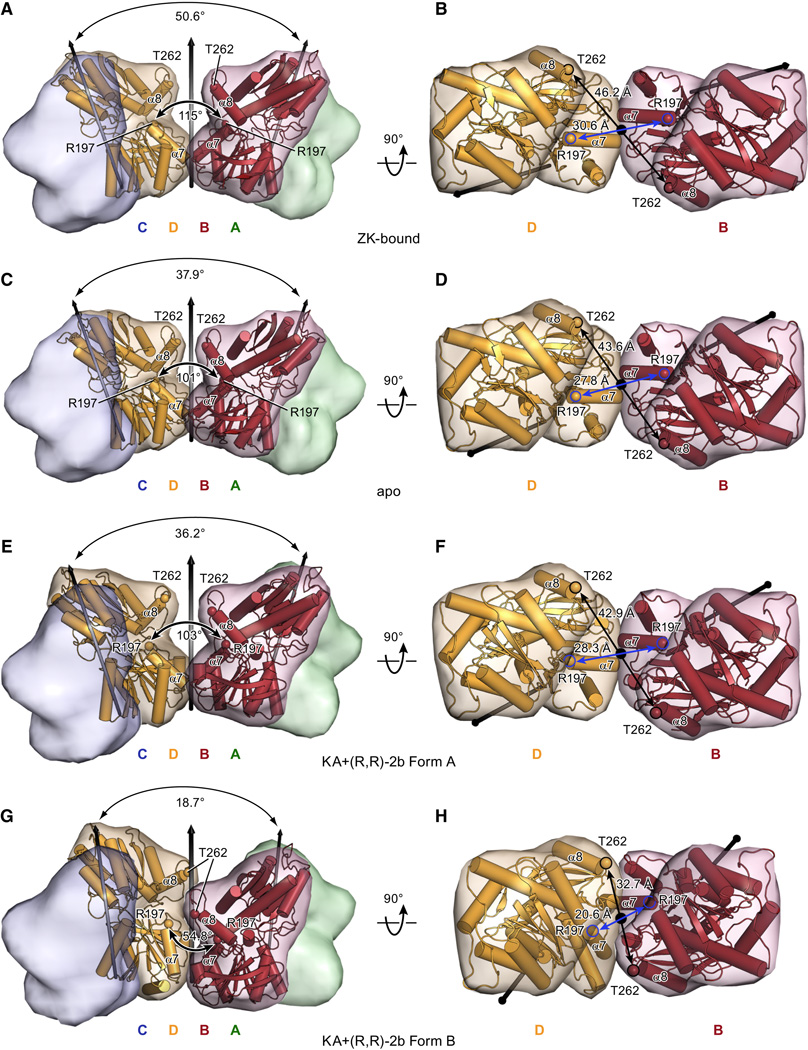
Ionotropic glutamate receptors (iGluRs) mediate the majority of fast excitatory signaling in the nervous system. Despite the profound importance of iGluRs to neurotransmission, little is known about the structures and dynamics of intact receptors in distinct functional states. Here, we elucidate the structures of the intact GluA2 AMPA receptor in an apo resting/closed state, in an activated/pre-open state bound with partial agonists and a positive allosteric modulator, and in a desensitized/closed state in complex with fluorowilliardiine. To probe the conformational properties of these states, we carried out double electron-electron resonance experiments on cysteine mutants and cryoelectron microscopy studies. We show how agonist binding modulates the conformation of the ligand-binding domain "layer" of the intact receptors and how, upon desensitization, the receptor undergoes large conformational rearrangements of the amino-terminal and ligand-binding domains. We define mechanistic principles by which to understand antagonism, activation, and desensitization in AMPA iGluRs.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Armstrong N, Gouaux E. Mechanisms for activation and antagonism of an AMPA-sensitive glutamate receptor: crystal structures of the GluR2 ligand binding core. Neuron. 2000;28:165–181. - PubMed
-
- Armstrong N, Jasti J, Beich-Frandsen M, Gouaux E. Measurement of conformational changes accompanying desensitization in an ionotropic glutamate receptor. Cell. 2006;127:85–97. - PubMed
-
- Bertolino M, Baraldi M, Parenti C, Braghiroli D, DiBella M, Vicini S, Costa E. Modulation of AMPA/kainate receptors by analogues of diazoxide and cyclothiazide in thin slices of rat hippocampus. Receptors & channels. 1993;1:267–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 1S10RR028832-01/RR/NCRR NIH HHS/United States
- R01 GM077659/GM/NIGMS NIH HHS/United States
- S10 RR027091/RR/NCRR NIH HHS/United States
- F32 MH100331/MH/NIMH NIH HHS/United States
- F32MH100331/MH/NIMH NIH HHS/United States
- U54 GM087519/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P30 NS061800/NS/NINDS NIH HHS/United States
- S10RR027091/RR/NCRR NIH HHS/United States
- S10 RR028832/RR/NCRR NIH HHS/United States
- R01 NS038631/NS/NINDS NIH HHS/United States
- R37 NS038631/NS/NINDS NIH HHS/United States
- U54-GM087519/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
