

Recognition and repair of chemically heterogeneous structures at DNA ends
- PMID: 25111769
- PMCID: PMC4303525
- DOI: 10.1002/em.21892
Recognition and repair of chemically heterogeneous structures at DNA ends
Abstract
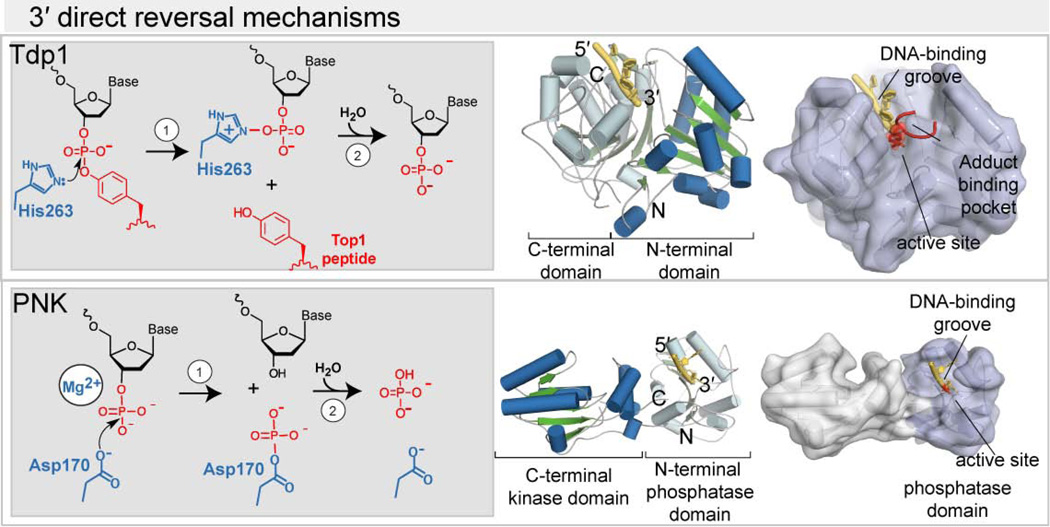
Exposure to environmental toxicants and stressors, radiation, pharmaceutical drugs, inflammation, cellular respiration, and routine DNA metabolism all lead to the production of cytotoxic DNA strand breaks. Akin to splintered wood, DNA breaks are not "clean." Rather, DNA breaks typically lack DNA 5'-phosphate and 3'-hydroxyl moieties required for DNA synthesis and DNA ligation. Failure to resolve damage at DNA ends can lead to abnormal DNA replication and repair, and is associated with genomic instability, mutagenesis, neurological disease, ageing and carcinogenesis. An array of chemically heterogeneous DNA termini arises from spontaneously generated DNA single-strand and double-strand breaks (SSBs and DSBs), and also from normal and/or inappropriate DNA metabolism by DNA polymerases, DNA ligases and topoisomerases. As a front line of defense to these genotoxic insults, eukaryotic cells have accrued an arsenal of enzymatic first responders that bind and protect damaged DNA termini, and enzymatically tailor DNA ends for DNA repair synthesis and ligation. These nucleic acid transactions employ direct damage reversal enzymes including Aprataxin (APTX), Polynucleotide kinase phosphatase (PNK), the tyrosyl DNA phosphodiesterases (TDP1 and TDP2), the Ku70/80 complex and DNA polymerase β (POLβ). Nucleolytic processing enzymes such as the MRE11/RAD50/NBS1/CtIP complex, Flap endonuclease (FEN1) and the apurinic endonucleases (APE1 and APE2) also act in the chemical "cleansing" of DNA breaks to prevent genomic instability and disease, and promote progression of DNA- and RNA-DNA damage response (DDR and RDDR) pathways. Here, we provide an overview of cellular first responders dedicated to the detection and repair of abnormal DNA termini.
Keywords: DNA damage response; X-ray crystallography; cancer; neurodegeneration.
© 2014 Wiley Periodicals, Inc.
Figures






References
-
- Ahel I, Rass U, El-Khamisy SF, Katyal S, Clements PM, McKinnon PJ, Caldecott KW, West SC. The neurodegenerative disease protein aprataxin resolves abortive DNA ligation intermediates. Nature. 2006;443:713–716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
