

Context-dependent control of alternative splicing by RNA-binding proteins
- PMID: 25112293
- PMCID: PMC4440546
- DOI: 10.1038/nrg3778
Context-dependent control of alternative splicing by RNA-binding proteins
Abstract
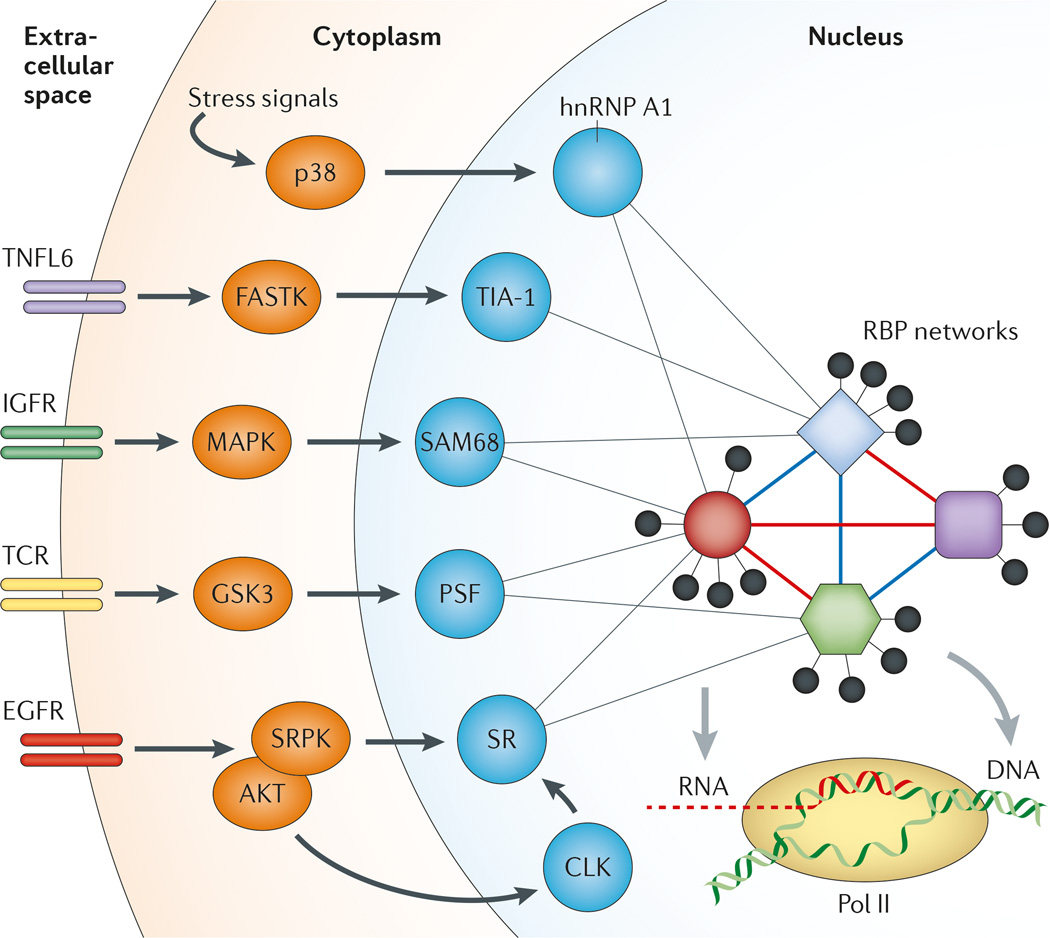
Sequence-specific RNA-binding proteins (RBPs) bind to pre-mRNA to control alternative splicing, but it is not yet possible to read the 'splicing code' that dictates splicing regulation on the basis of genome sequence. Each alternative splicing event is controlled by multiple RBPs, the combined action of which creates a distribution of alternatively spliced products in a given cell type. As each cell type expresses a distinct array of RBPs, the interpretation of regulatory information on a given RNA target is exceedingly dependent on the cell type. RBPs also control each other's functions at many levels, including by mutual modulation of their binding activities on specific regulatory RNA elements. In this Review, we describe some of the emerging rules that govern the highly context-dependent and combinatorial nature of alternative splicing regulation.
Figures
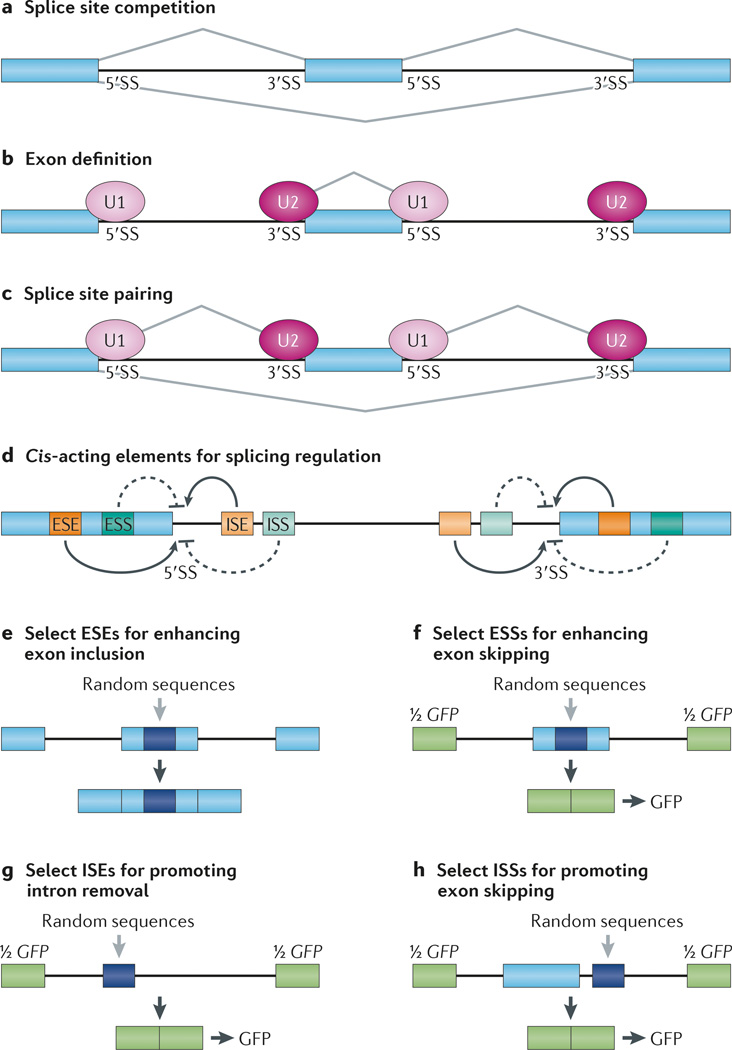
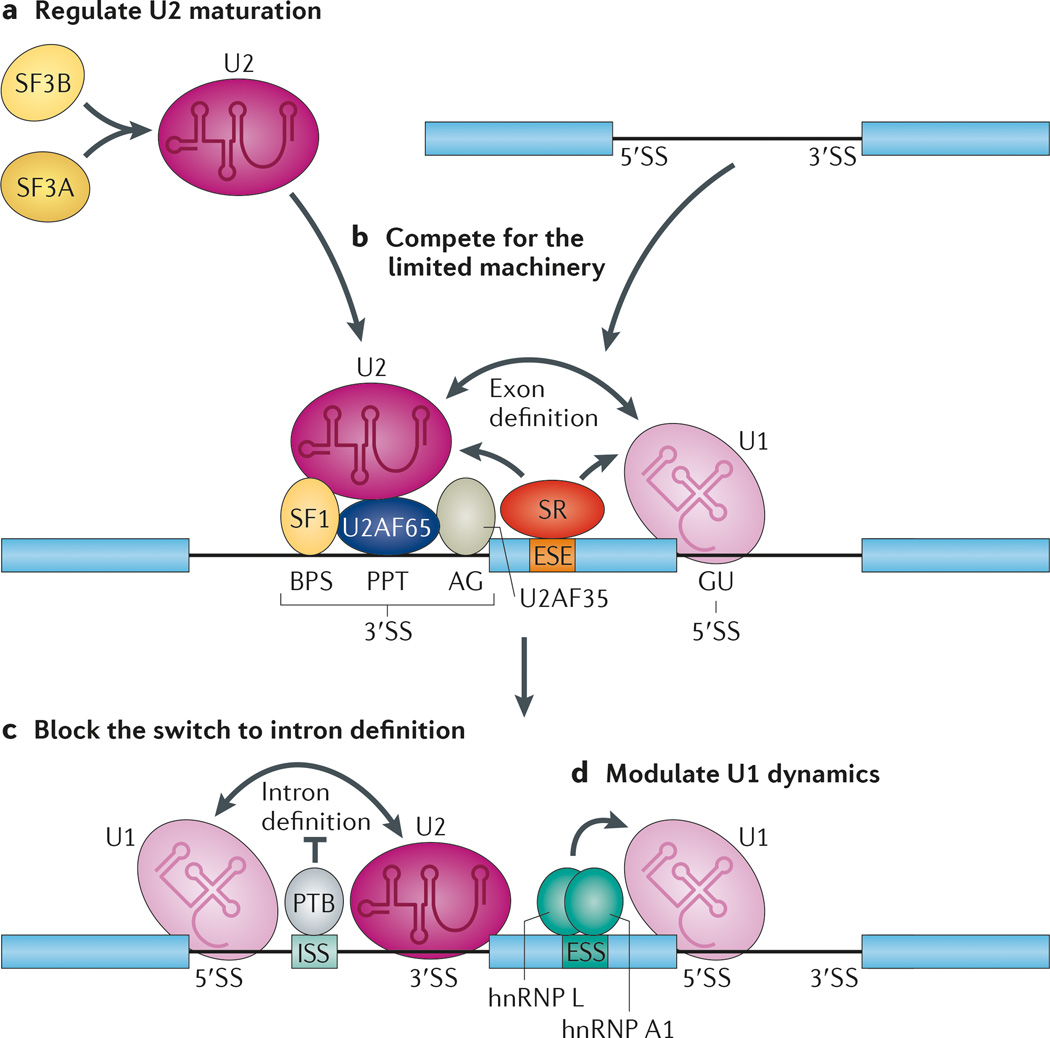
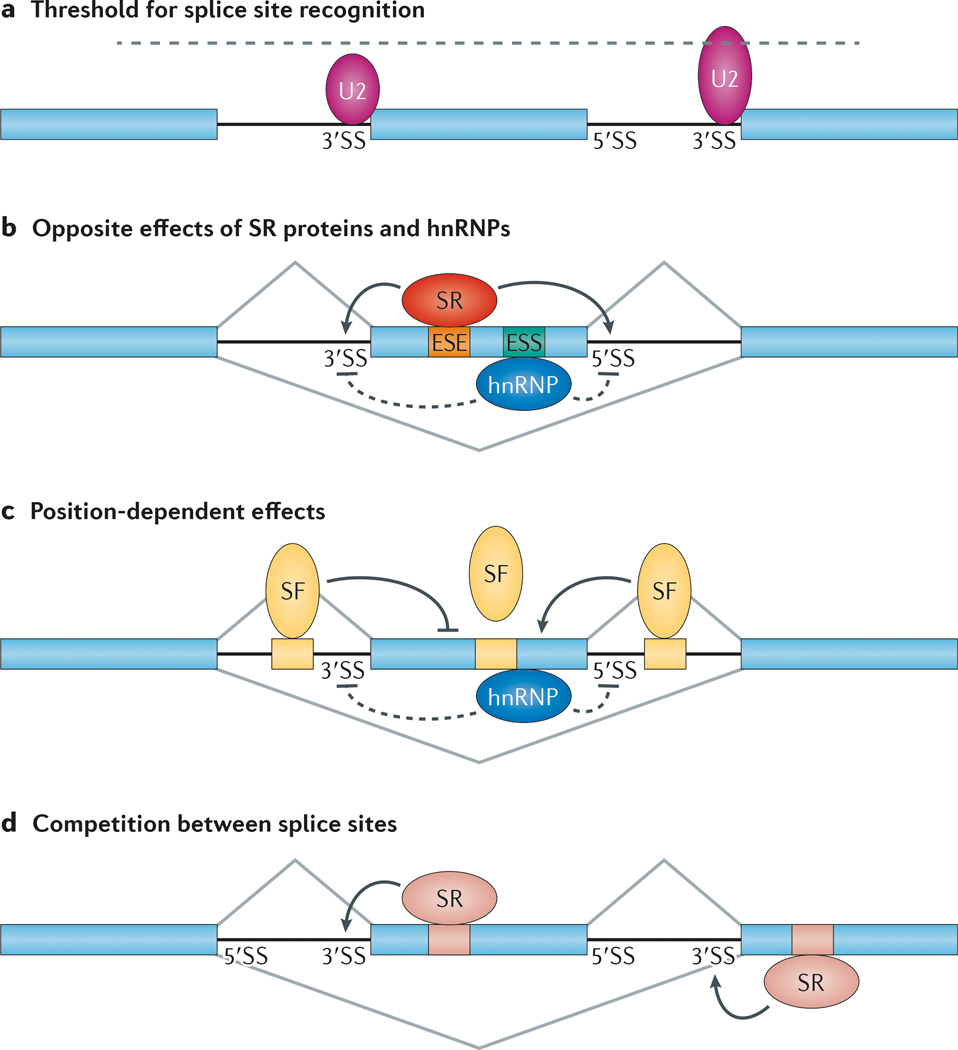
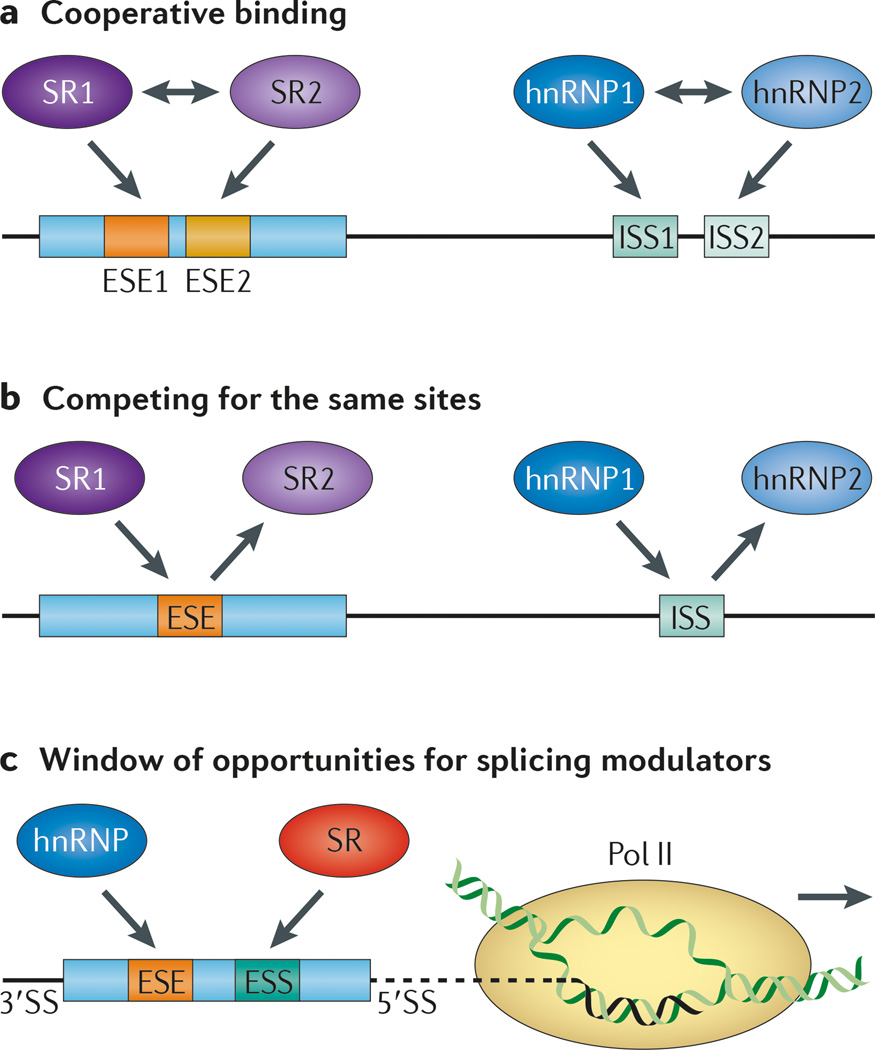
References
-
- Penny D, Hoeppner MP, Poole AM, Jeffares DC. An overview of the introns-first theory. J. Mol. Evol. 2009;69:527–540. - PubMed
-
- Ast G. How did alternative splicing evolve? Nature Rev. Genet. 2004;5:773–782. - PubMed
-
- Keren H, Lev-Maor G, Ast G. Alternative splicing and evolution: diversification, exon definition and function. Nature Rev. Genet. 2010;11:345–355. - PubMed
-
- Brown JB, et al. Diversity and dynamics of the Drosophila transcriptome. Nature. 2014 http://dx.doi. org/10.1038/nature12962. - DOI - PMC - PubMed
-
- Barbosa-Morais NL, et al. The evolutionary landscape of alternative splicing in vertebrate species. Science. 2012;338:1587–1593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
