

A distinct immunogenic region of glutamic acid decarboxylase 65 is naturally processed and presented by human islet cells to cytotoxic CD8 T cells
- PMID: 25112375
- PMCID: PMC4260902
- DOI: 10.1111/cei.12436
A distinct immunogenic region of glutamic acid decarboxylase 65 is naturally processed and presented by human islet cells to cytotoxic CD8 T cells
Abstract
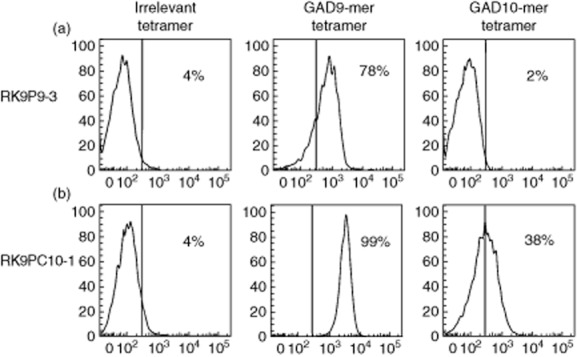
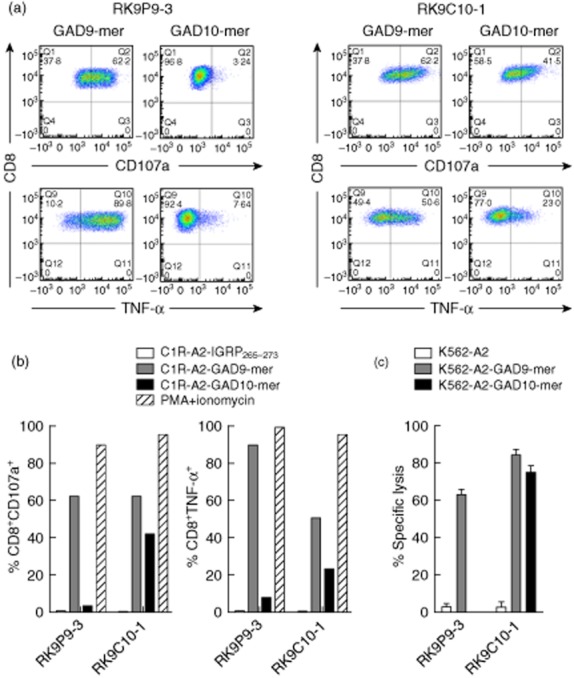
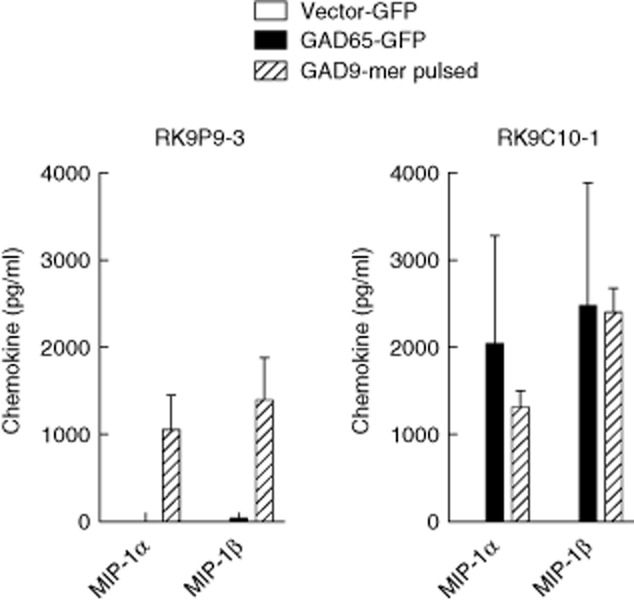
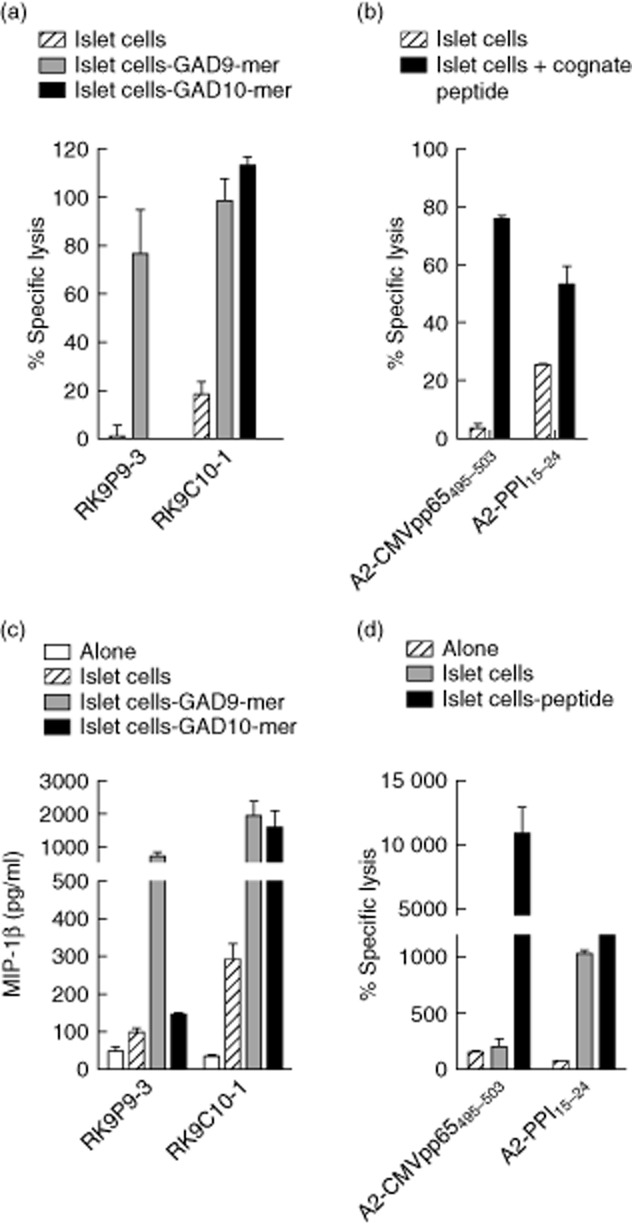
CD8 T cells specific for islet autoantigens are major effectors of β cell damage in type 1 diabetes, and measurement of their number and functional characteristics in blood represent potentially important disease biomarkers. CD8 T cell reactivity against glutamic acid decarboxylase 65 (GAD65) in HLA-A*0201 subjects has been reported to focus on an immunogenic region 114-123 (VMNILLQYVV), with studies demonstrating both 114-123 and 114-122 epitopes being targeted. However, the fine specificity of this response is unclear and the key question as to which epitope(s) β cells naturally process and present and, therefore, the pathogenic potential of CD8 T cells with different specificities within this region has not been addressed. We generated human leucocyte antigen (HLA)-A*0201-restricted CD8 T cell clones recognizing either 114-122 alone or both 114-122 and 114-123. Both clone types show potent and comparable effector functions (cytokine and chemokine secretion) and killing of indicator target cells externally pulsed with cognate peptide. However, only clones recognizing 114-123 kill target cells transfected with HLA-A*0201 and GAD2 and HLA-A*0201(+) human islet cells. We conclude that the endogenous pathway of antigen processing by HLA-A*0201-expressing cells generates GAD65114-123 as the predominant epitope in this region. These studies highlight the importance of understanding β cell epitope presentation in the design of immune monitoring for potentially pathogenic CD8 T cells.
Keywords: CD8 T cell clones; GAD65; autoimmunity; peptide-processing; type 1 diabetes.
© 2014 British Society for Immunology.
Figures
Similar articles
-
Dendritic Cells Guide Islet Autoimmunity through a Restricted and Uniquely Processed Peptidome Presented by High-Risk HLA-DR.J Immunol. 2016 Apr 15;196(8):3253-63. doi: 10.4049/jimmunol.1501282. Epub 2016 Mar 4. J Immunol. 2016. PMID: 26944932
-
Identification of a naturally processed cytotoxic CD8 T-cell epitope of coxsackievirus B4, presented by HLA-A2.1 and located in the PEVKEK region of the P2C nonstructural protein.J Virol. 2004 Dec;78(24):13399-408. doi: 10.1128/JVI.78.24.13399-13408.2004. J Virol. 2004. PMID: 15564450 Free PMC article.
-
Evidence for molecular mimicry between human T cell epitopes in rotavirus and pancreatic islet autoantigens.J Immunol. 2010 Feb 15;184(4):2204-10. doi: 10.4049/jimmunol.0900709. Epub 2010 Jan 18. J Immunol. 2010. PMID: 20083660
-
Diabetogenic T-cell clones.Diabetes. 1996 Oct;45(10):1299-305. doi: 10.2337/diab.45.10.1299. Diabetes. 1996. PMID: 8826963 Review.
-
Interactions of effectors and regulators are decisive in the manifestations of type 1 diabetes in nonobese diabetic mice.Curr Dir Autoimmun. 2001;4:171-92. doi: 10.1159/000060537. Curr Dir Autoimmun. 2001. PMID: 11569402 Review. No abstract available.
Cited by
-
Thermal Stability of Heterotrimeric pMHC Proteins as Determined by Circular Dichroism Spectroscopy.Bio Protoc. 2017 Jul 5;7(13):e2366. doi: 10.21769/BioProtoc.2366. Bio Protoc. 2017. PMID: 28748203 Free PMC article.
-
Autoreactive T effector memory differentiation mirrors β cell function in type 1 diabetes.J Clin Invest. 2018 Aug 1;128(8):3460-3474. doi: 10.1172/JCI120555. Epub 2018 Jul 16. J Clin Invest. 2018. PMID: 29851415 Free PMC article. Clinical Trial.
-
T cell epitopes and post-translationally modified epitopes in type 1 diabetes.Curr Diab Rep. 2015 Nov;15(11):90. doi: 10.1007/s11892-015-0657-7. Curr Diab Rep. 2015. PMID: 26370701 Free PMC article. Review.
-
T cells in the control of organ-specific autoimmunity.J Clin Invest. 2015 Jun;125(6):2250-60. doi: 10.1172/JCI78089. Epub 2015 May 18. J Clin Invest. 2015. PMID: 25985270 Free PMC article. Review.
-
Optimized Peptide-MHC Multimer Protocols for Detection and Isolation of Autoimmune T-Cells.Front Immunol. 2018 Jun 29;9:1378. doi: 10.3389/fimmu.2018.01378. eCollection 2018. Front Immunol. 2018. PMID: 30008714 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
