

Evidence for a molecular diode-based mechanism in a multispecific ATP-binding cassette (ABC) exporter: SER-1368 as a gatekeeping residue in the yeast multidrug transporter Pdr5
- PMID: 25112867
- PMCID: PMC4176199
- DOI: 10.1074/jbc.M114.586032
Evidence for a molecular diode-based mechanism in a multispecific ATP-binding cassette (ABC) exporter: SER-1368 as a gatekeeping residue in the yeast multidrug transporter Pdr5
Abstract
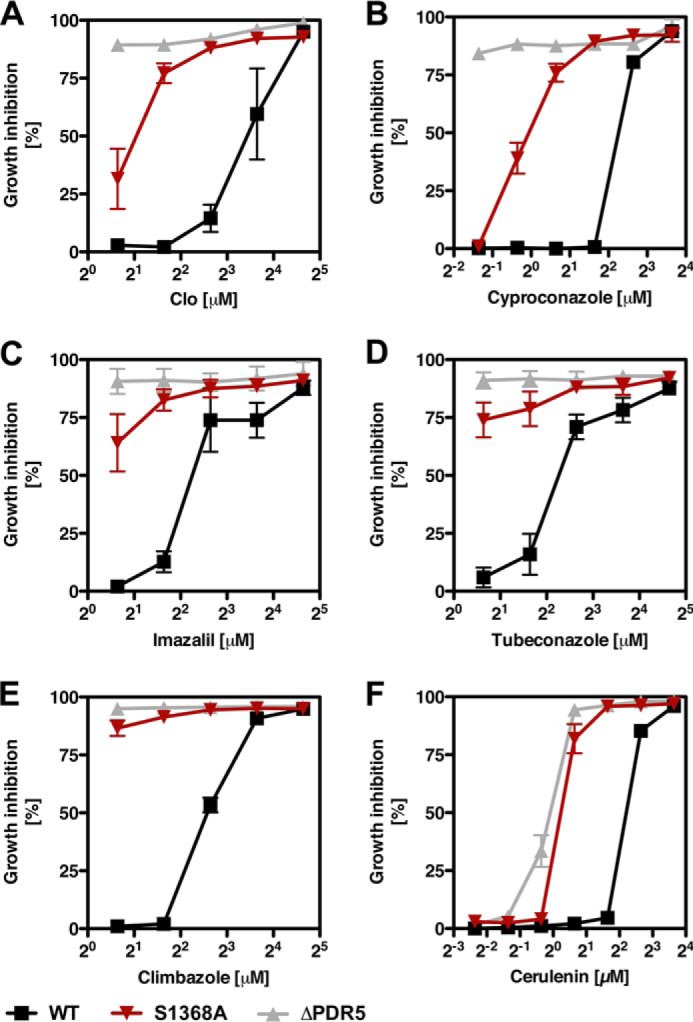
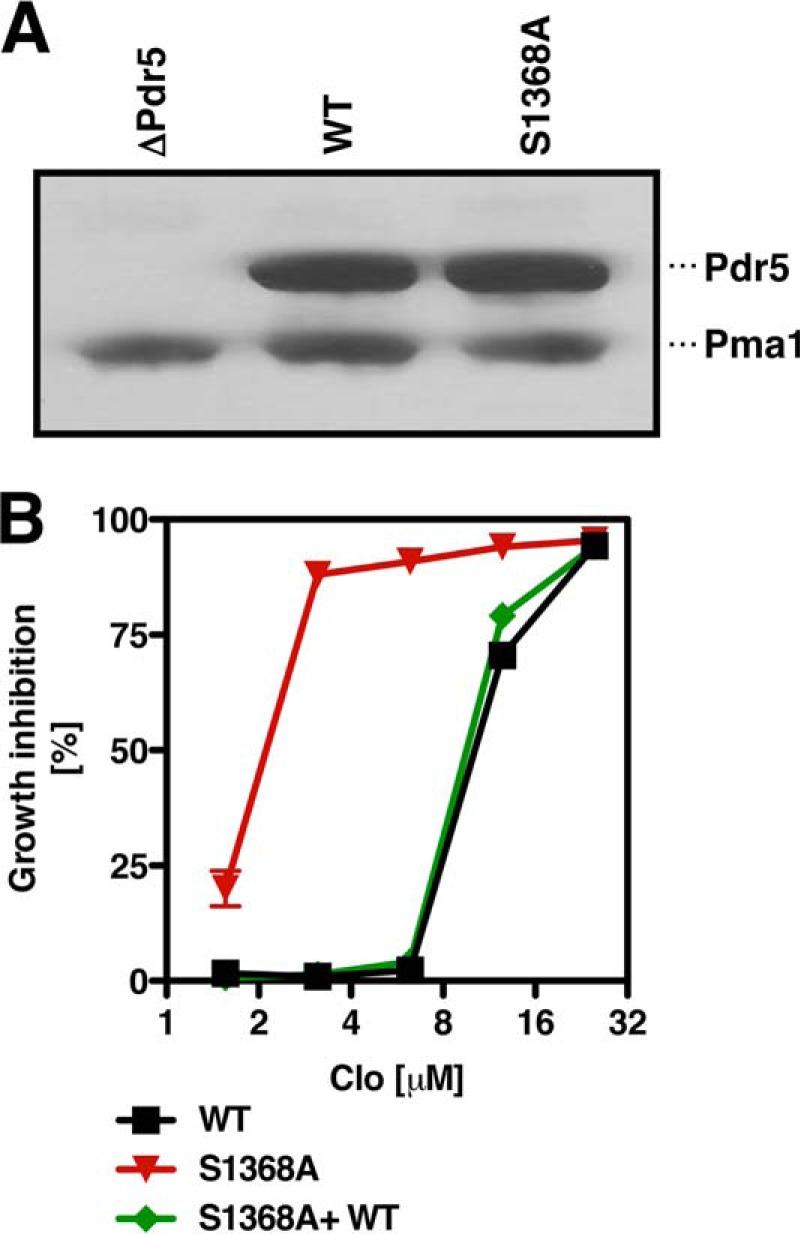
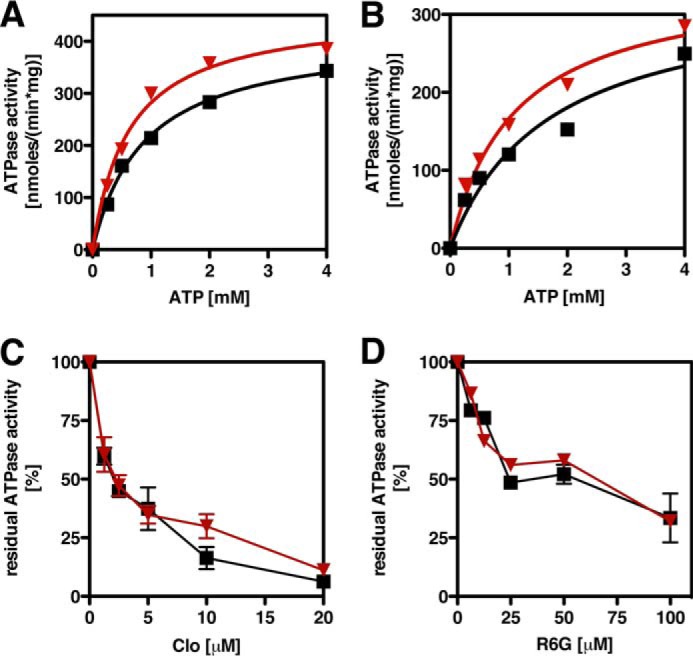
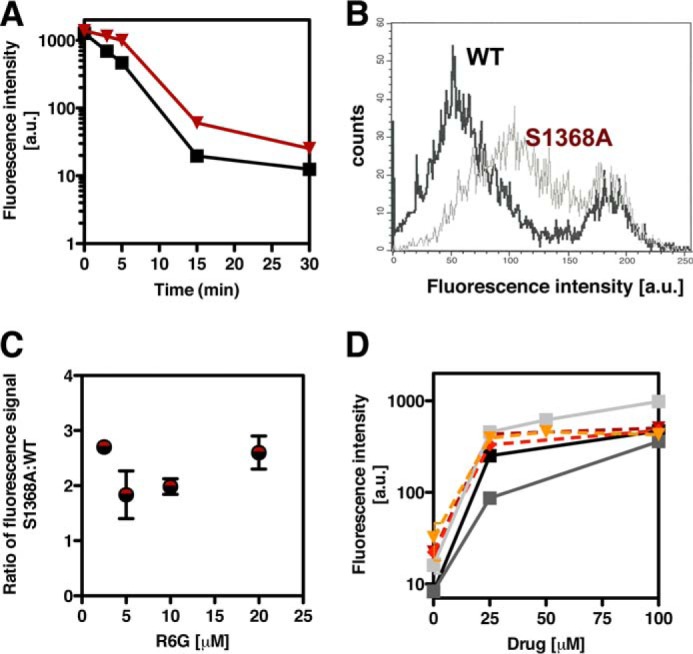
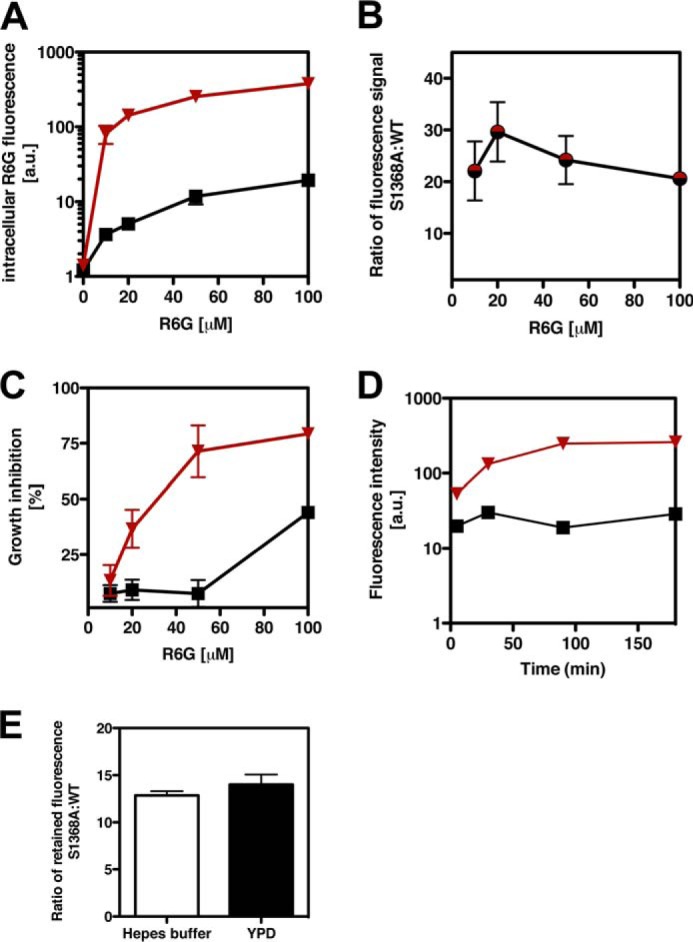
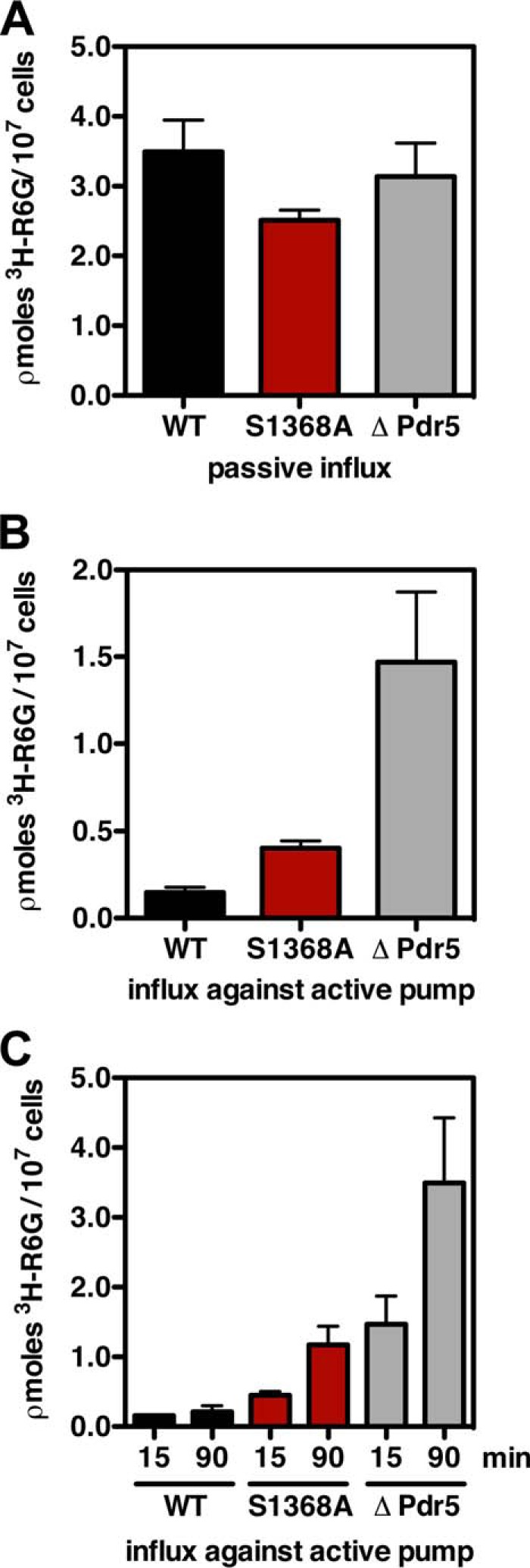
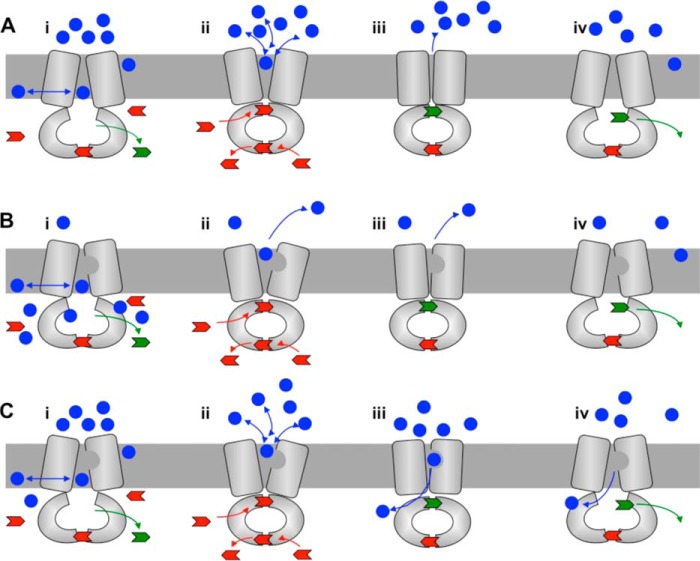
ATP-binding cassette multidrug efflux pumps transport a wide range of substrates. Current models suggest that a drug binds relatively tightly to a transport site in the transmembrane domains when the protein is in the closed inward facing conformation. Upon binding of ATP, the transporter can switch to an outward facing (drug off or drug releasing) structure of lower affinity. ATP hydrolysis is critically important for remodeling the drug-binding site to facilitate drug release and to reset the transporter for a new transport cycle. We characterized the novel phenotype of an S1368A mutant that lies in the putative drug-binding pocket of the yeast multidrug transporter Pdr5. This substitution created broad, severe drug hypersensitivity, although drug binding, ATP hydrolysis, and intradomain signaling were indistinguishable from the wild-type control. Several different rhodamine 6G efflux and accumulation assays yielded evidence consistent with the possibility that Ser-1368 prevents reentry of the excluded drug.
Keywords: ABC Transporter; ATPase; Drug Transport; Membrane Protein; Multidrug Transporter.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Gottesman M.M., Fojo T., Bates S. E. (2002) Multidrug resistance in cancer: role of ATP-dependent transporters. Nat. Rev. Cancer 2, 48–58 - PubMed
-
- Kontoyiannis D. P., Lewis R. E. (2002) Antifungal drug resistance of pathogenic fungi. Lancet 359, 1135–1144 - PubMed
-
- Monk B. C., Goffeau A. (2008) Multidrug resistance to antifungals. Science 321, 367–369 - PubMed
-
- Dawson R. J., Locher K. P. (2006) Structure of a bacterial multidrug ABC transporter. Nature 443, 180–185 - PubMed
-
- Oldham M. L., Khare D., Quiocho F. A., Davidson A. L., Chen J. (2007) Crystal structure of a catalytic intermediate of the maltose transporter. Nature 450, 515–521 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
