

Macromolecular transport between the nucleus and the cytoplasm: Advances in mechanism and emerging links to disease
- PMID: 25116306
- PMCID: PMC4161953
- DOI: 10.1016/j.bbamcr.2014.08.003
Macromolecular transport between the nucleus and the cytoplasm: Advances in mechanism and emerging links to disease
Abstract
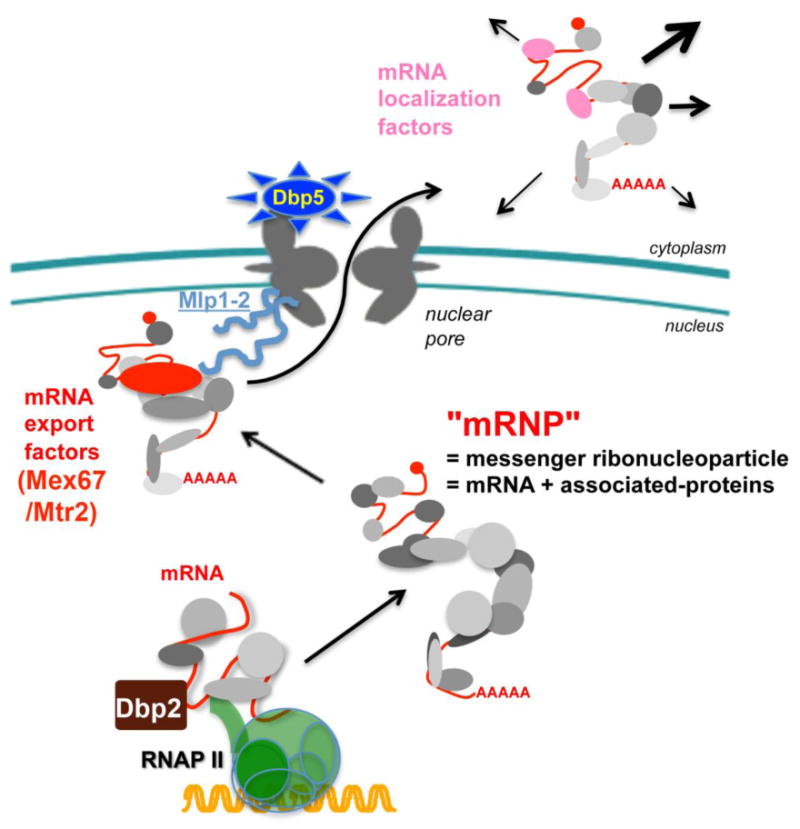
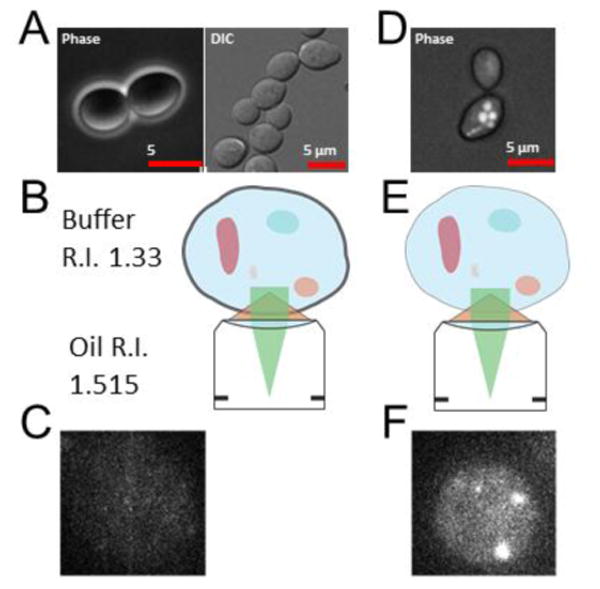
Transport of macromolecules between the cytoplasm and the nucleus is critical for the function of all eukaryotic cells. Large macromolecular channels termed nuclear pore complexes that span the nuclear envelope mediate the bidirectional transport of cargoes between the nucleus and cytoplasm. However, the influence of macromolecular trafficking extends past the nuclear pore complex to transcription and RNA processing within the nucleus and signaling pathways that reach into the cytoplasm and beyond. At the Mechanisms of Nuclear Transport biennial meeting held from October 18 to 23, 2013 in Woods Hole, MA, researchers in the field met to report on their recent findings. The work presented highlighted significant advances in understanding nucleocytoplasmic trafficking including how transport receptors and cargoes pass through the nuclear pore complex, the many signaling pathways that impinge on transport pathways, interplay between the nuclear envelope, nuclear pore complexes, and transport pathways, and numerous links between transport pathways and human disease. The goal of this review is to highlight newly emerging themes in nuclear transport and underscore the major questions that are likely to be the focus of future research in the field.
Keywords: Karyopherin/importin/exportin; Nuclear pore; Nucleocytoplasmic transport; Protein import; RNA export; RNA processing.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Karyopherin-mediated nucleocytoplasmic transport.Nat Rev Mol Cell Biol. 2022 May;23(5):307-328. doi: 10.1038/s41580-021-00446-7. Epub 2022 Jan 20. Nat Rev Mol Cell Biol. 2022. PMID: 35058649 Free PMC article. Review.
-
On the asymmetric partitioning of nucleocytoplasmic transport - recent insights and open questions.J Cell Sci. 2021 Apr 1;134(7):jcs240382. doi: 10.1242/jcs.240382. Epub 2021 Apr 13. J Cell Sci. 2021. PMID: 33912945 Review.
-
The nuclear pore complex: mediator of translocation between nucleus and cytoplasm.J Cell Sci. 2000 May;113 ( Pt 10):1651-9. doi: 10.1242/jcs.113.10.1651. J Cell Sci. 2000. PMID: 10769196 Review.
-
Nucleocytoplasmic Shuttling of Porcine Parvovirus NS1 Protein Mediated by the CRM1 Nuclear Export Pathway and the Importin α/β Nuclear Import Pathway.J Virol. 2022 Jan 12;96(1):e0148121. doi: 10.1128/JVI.01481-21. Epub 2021 Oct 13. J Virol. 2022. PMID: 34643426 Free PMC article.
-
Nucleocytoplasmic transport of macromolecules.Microbiol Mol Biol Rev. 1997 Jun;61(2):193-211. doi: 10.1128/mmbr.61.2.193-211.1997. Microbiol Mol Biol Rev. 1997. PMID: 9184010 Free PMC article. Review.
Cited by
-
Nuclear export protein CSE1L interacts with P65 and promotes NSCLC growth via NF-κB/MAPK pathway.Mol Ther Oncolytics. 2021 Mar 10;21:23-36. doi: 10.1016/j.omto.2021.02.015. eCollection 2021 Jun 25. Mol Ther Oncolytics. 2021. PMID: 33869740 Free PMC article.
-
IPO5 promotes the proliferation and tumourigenicity of colorectal cancer cells by mediating RASAL2 nuclear transportation.J Exp Clin Cancer Res. 2019 Jul 9;38(1):296. doi: 10.1186/s13046-019-1290-0. J Exp Clin Cancer Res. 2019. PMID: 31288861 Free PMC article.
-
Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis.Int J Mol Sci. 2021 Jun 30;22(13):7074. doi: 10.3390/ijms22137074. Int J Mol Sci. 2021. PMID: 34209205 Free PMC article. Review.
-
Thyroid hormone receptor localization in target tissues.J Endocrinol. 2018 Apr;237(1):R19-R34. doi: 10.1530/JOE-17-0708. Epub 2018 Feb 12. J Endocrinol. 2018. PMID: 29440347 Free PMC article. Review.
-
Recognition motifs for importin 4 [(L)PPRS(G/P)P] and importin 5 [KP(K/Y)LV] binding, identified by bio-informatic simulation and experimental in vitro validation.Comput Struct Biotechnol J. 2022 Oct 26;20:5952-5961. doi: 10.1016/j.csbj.2022.10.015. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36382187 Free PMC article.
References
-
- Raices M, D'Angelo MA. Nuclear pore complex composition: a new regulator of tissue-specific and developmental functions. Nature reviews Molecular cell biology. 2012;13:687–699. - PubMed
-
- Grossman E, Medalia O, Zwerger M. Functional architecture of the nuclear pore complex. Annual review of biophysics. 2012;41:557–584. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
