

The transcription factor NRSF contributes to epileptogenesis by selective repression of a subset of target genes
- PMID: 25117540
- PMCID: PMC4129437
- DOI: 10.7554/eLife.01267
The transcription factor NRSF contributes to epileptogenesis by selective repression of a subset of target genes
Abstract
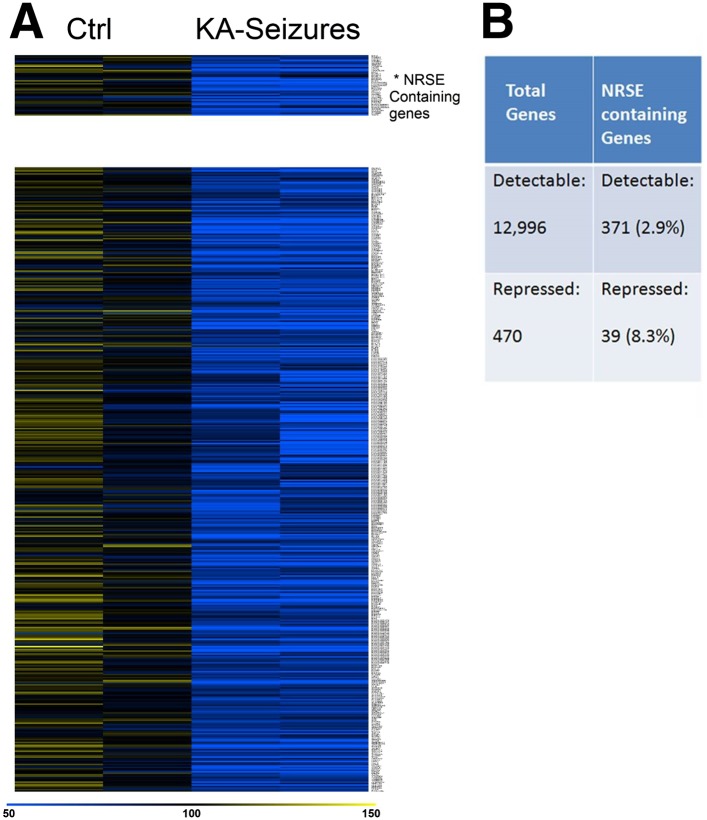
The mechanisms generating epileptic neuronal networks following insults such as severe seizures are unknown. We have previously shown that interfering with the function of the neuron-restrictive silencer factor (NRSF/REST), an important transcription factor that influences neuronal phenotype, attenuated development of this disorder. In this study, we found that epilepsy-provoking seizures increased the low NRSF levels in mature hippocampus several fold yet surprisingly, provoked repression of only a subset (∼10%) of potential NRSF target genes. Accordingly, the repressed gene-set was rescued when NRSF binding to chromatin was blocked. Unexpectedly, genes selectively repressed by NRSF had mid-range binding frequencies to the repressor, a property that rendered them sensitive to moderate fluctuations of NRSF levels. Genes selectively regulated by NRSF during epileptogenesis coded for ion channels, receptors, and other crucial contributors to neuronal function. Thus, dynamic, selective regulation of NRSF target genes may play a role in influencing neuronal properties in pathological and physiological contexts.
Keywords: epilepsy; gene set enrichment analysis; neuron-restrictive silencing factor.
Copyright © 2014, McClelland et al.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures







Similar articles
-
Brain REST/NRSF Is Not Only a Silent Repressor but Also an Active Protector.Mol Neurobiol. 2017 Jan;54(1):541-550. doi: 10.1007/s12035-015-9658-4. Epub 2016 Jan 7. Mol Neurobiol. 2017. PMID: 26742529 Review.
-
Inhibition of Neuron-Restrictive Silencing Factor (REST/NRSF) Chromatin Binding Attenuates Epileptogenesis.eNeuro. 2024 May 17;11(5):ENEURO.0006-24.2024. doi: 10.1523/ENEURO.0006-24.2024. Print 2024 May. eNeuro. 2024. PMID: 38641413 Free PMC article.
-
Dual and Opposing Roles of MicroRNA-124 in Epilepsy Are Mediated through Inflammatory and NRSF-Dependent Gene Networks.Cell Rep. 2016 Mar 15;14(10):2402-12. doi: 10.1016/j.celrep.2016.02.042. Epub 2016 Mar 3. Cell Rep. 2016. PMID: 26947066 Free PMC article.
-
Neuronal expression of zinc finger transcription factor REST/NRSF/XBR gene.J Neurosci. 1998 Feb 15;18(4):1280-96. doi: 10.1523/JNEUROSCI.18-04-01280.1998. J Neurosci. 1998. PMID: 9454838 Free PMC article.
-
Regulation of the cholinergic gene locus by the repressor element-1 silencing transcription factor/neuron restrictive silencer factor (REST/NRSF).Life Sci. 2004 Mar 19;74(18):2213-25. doi: 10.1016/j.lfs.2003.08.045. Life Sci. 2004. PMID: 15017977 Review.
Cited by
-
NRSF and Its Epigenetic Effectors: New Treatments for Neurological Disease.Brain Sci. 2018 Dec 19;8(12):226. doi: 10.3390/brainsci8120226. Brain Sci. 2018. PMID: 30572571 Free PMC article. Review.
-
Applications for Deep Learning in Epilepsy Genetic Research.Int J Mol Sci. 2023 Sep 27;24(19):14645. doi: 10.3390/ijms241914645. Int J Mol Sci. 2023. PMID: 37834093 Free PMC article. Review.
-
Calcium Channel Subunit α2δ4 Is Regulated by Early Growth Response 1 and Facilitates Epileptogenesis.J Neurosci. 2019 Apr 24;39(17):3175-3187. doi: 10.1523/JNEUROSCI.1731-18.2019. Epub 2019 Feb 21. J Neurosci. 2019. PMID: 30792272 Free PMC article.
-
Brain REST/NRSF Is Not Only a Silent Repressor but Also an Active Protector.Mol Neurobiol. 2017 Jan;54(1):541-550. doi: 10.1007/s12035-015-9658-4. Epub 2016 Jan 7. Mol Neurobiol. 2017. PMID: 26742529 Review.
-
Rare and common epilepsies converge on a shared gene regulatory network providing opportunities for novel antiepileptic drug discovery.Genome Biol. 2016 Dec 13;17(1):245. doi: 10.1186/s13059-016-1097-7. Genome Biol. 2016. PMID: 27955713 Free PMC article.
References
-
- Andres ME, Burger C, Peral-Rubio MJ, Battaglioli E, Anderson ME, Grimes J, Dallman J, Ballas N, Mandel G. 1999. CoREST: a functional corepressor required for regulation of neural specific gene expression. Proceedings of the National Academy of Sciences of USA 96:9873–9878. doi: 10.1073/pnas.96.17.9873 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
