

Bidirectional cargo transport: moving beyond tug of war
- PMID: 25118718
- PMCID: PMC5014371
- DOI: 10.1038/nrm3853
Bidirectional cargo transport: moving beyond tug of war
Abstract
Vesicles, organelles and other intracellular cargo are transported by kinesin and dynein motors, which move in opposite directions along microtubules. This bidirectional cargo movement is frequently described as a 'tug of war' between oppositely directed molecular motors attached to the same cargo. However, although many experimental and modelling studies support the tug-of-war paradigm, numerous knockout and inhibition studies in various systems have found that inhibiting one motor leads to diminished motility in both directions, which is a 'paradox of co-dependence' that challenges the paradigm. In an effort to resolve this paradox, three classes of bidirectional transport models--microtubule tethering, mechanical activation and steric disinhibition--are proposed, and a general mathematical modelling framework for bidirectional cargo transport is put forward to guide future experiments.
Figures
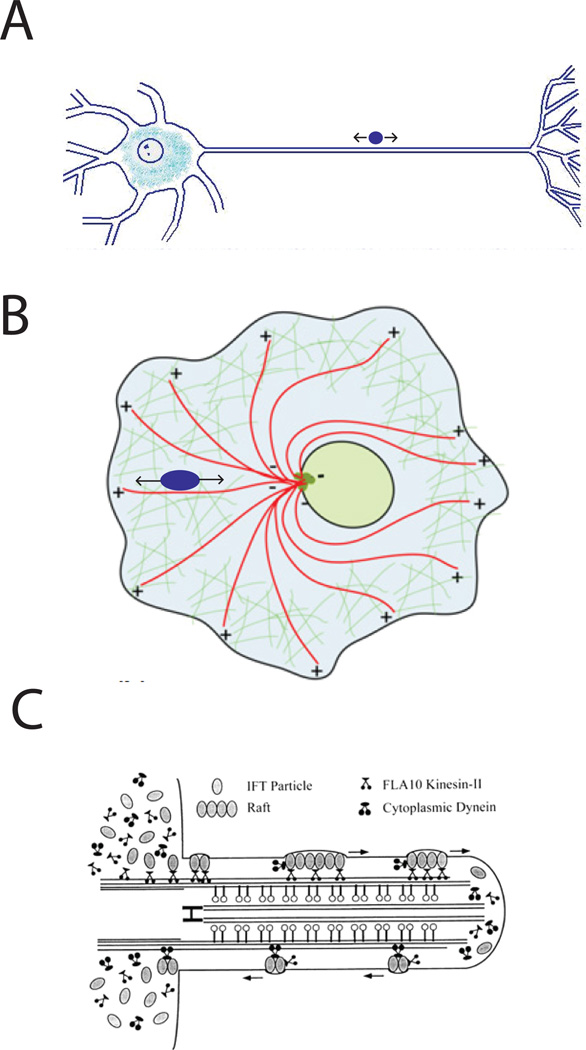
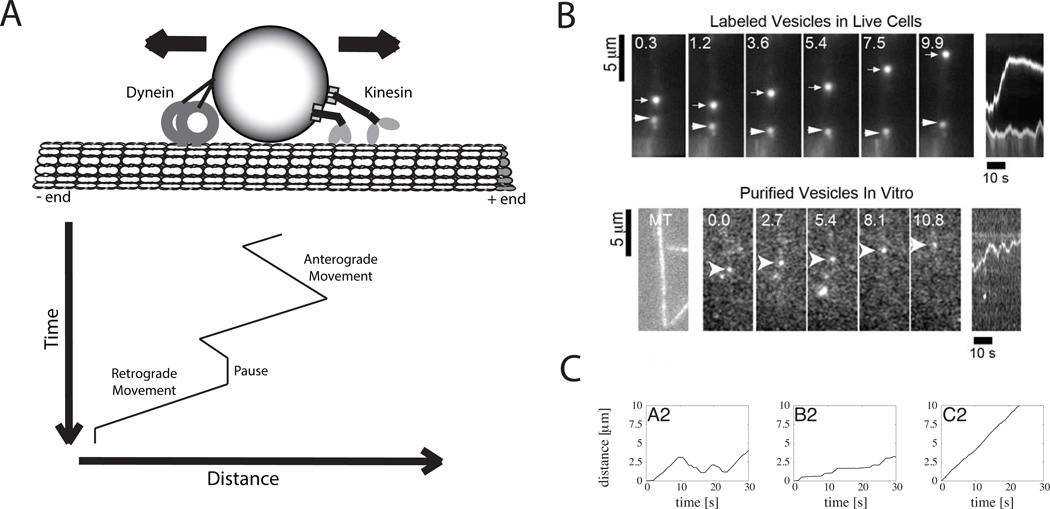
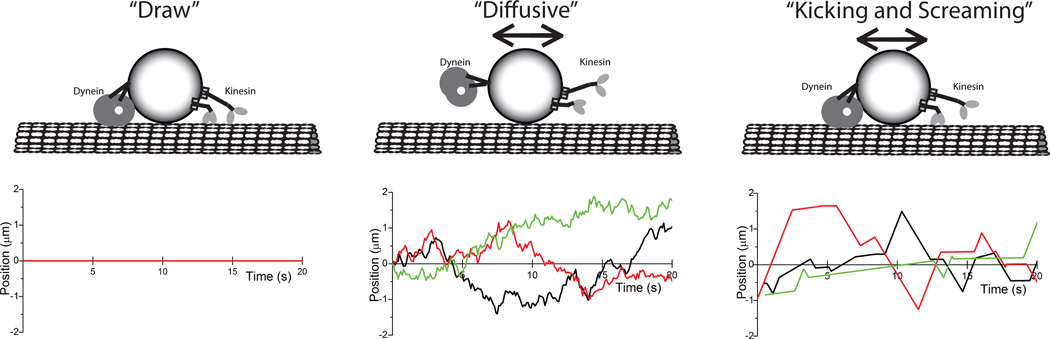
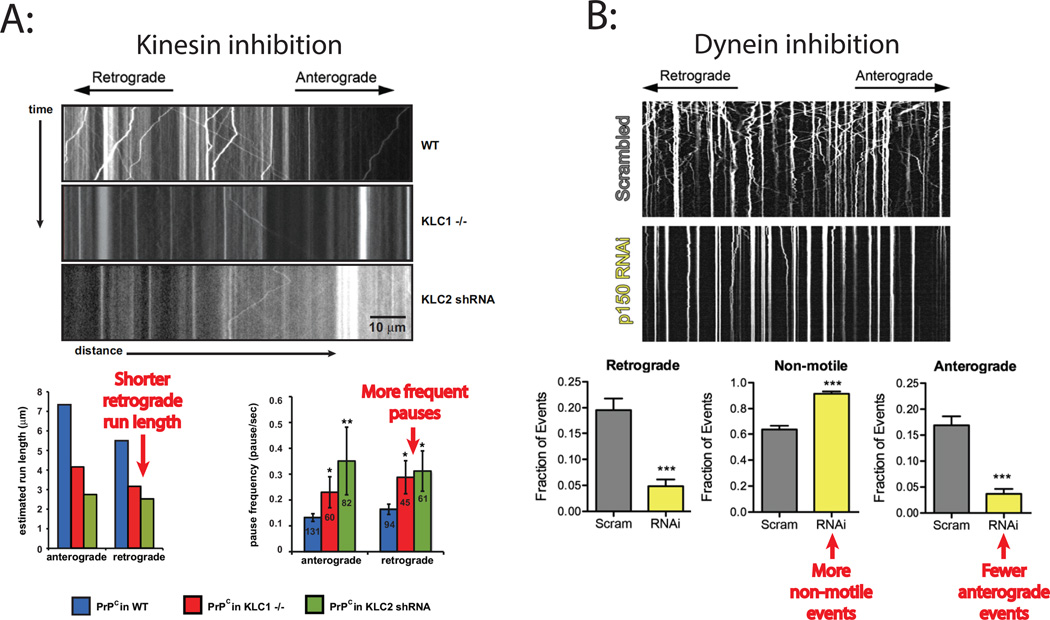
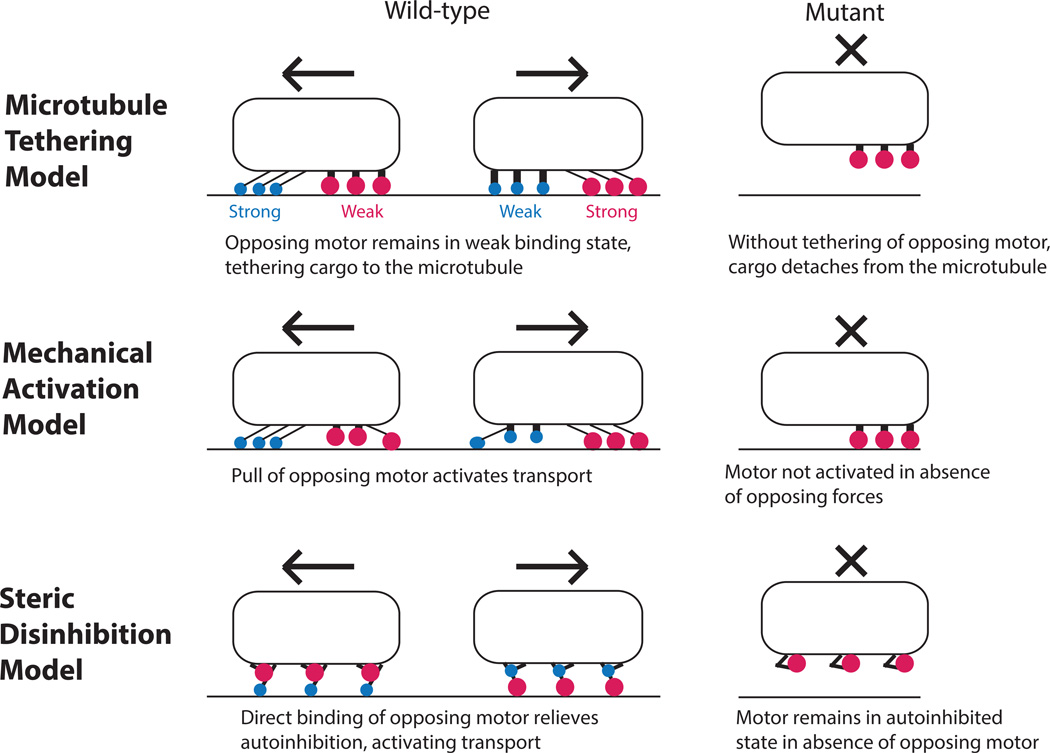
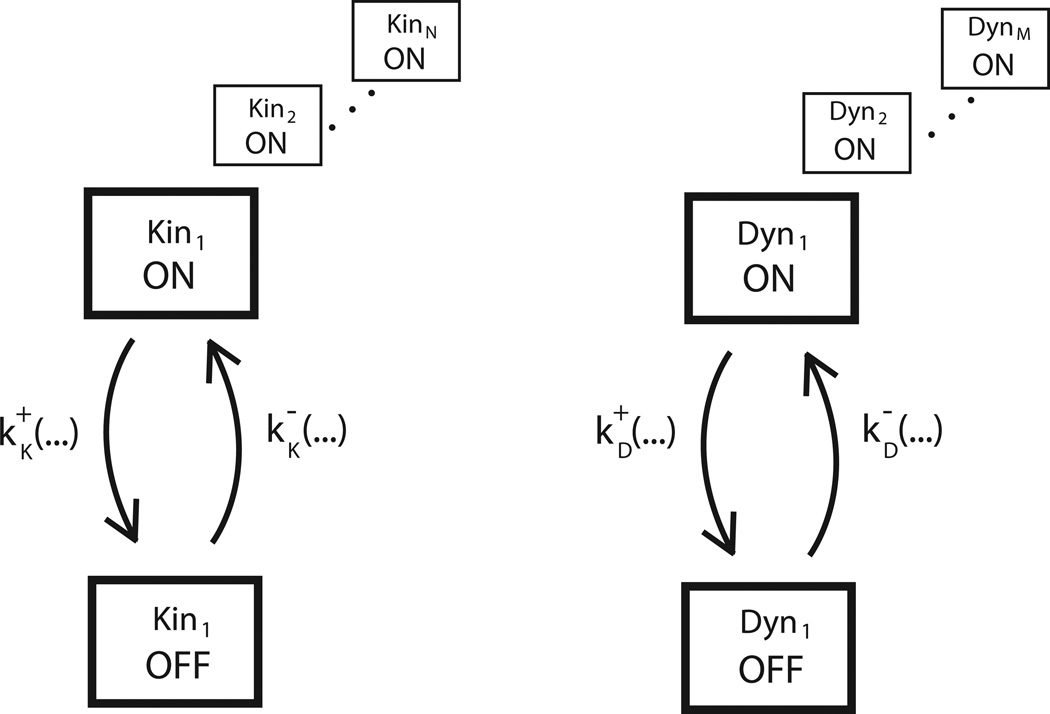
References
-
- Hirokawa N, Niwa S, Tanaka Y. Molecular motors in neurons: transport mechanisms and roles in brain function, development, and disease. Neuron. 2010;68:610–638. - PubMed
-
- Hirokawa N, Noda Y, Tanaka Y, Niwa S. Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Biol. 2009;10:682–696. - PubMed
-
- Ma S, Chisholm RL. Cytoplasmic dynein-associated structures move bidirectionally in vivo. J Cell Sci. 2002;115:1453–1460. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
