

Differential glycosylation of envelope gp120 is associated with differential recognition of HIV-1 by virus-specific antibodies and cell infection
- PMID: 25120578
- PMCID: PMC4130436
- DOI: 10.1186/1742-6405-11-23
Differential glycosylation of envelope gp120 is associated with differential recognition of HIV-1 by virus-specific antibodies and cell infection
Abstract
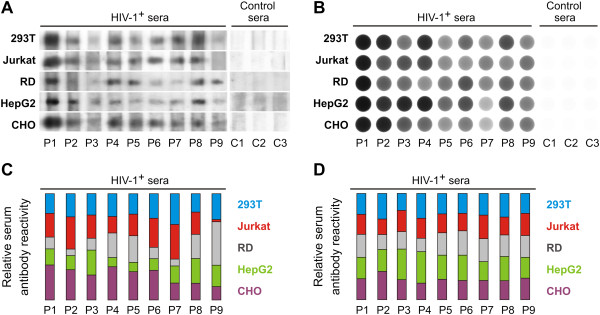
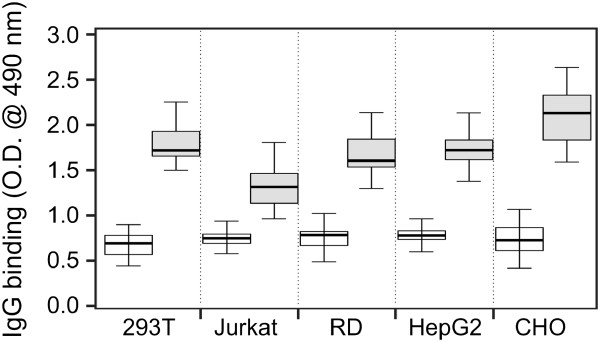
Background: HIV-1 entry into host cells is mediated by interactions between the virus envelope glycoprotein (gp120/gp41) and host-cell receptors. N-glycans represent approximately 50% of the molecular mass of gp120 and serve as potential antigenic determinants and/or as a shield against immune recognition. We previously reported that N-glycosylation of recombinant gp120 varied, depending on the producer cells, and the glycosylation variability affected gp120 recognition by serum antibodies from persons infected with HIV-1 subtype B. However, the impact of gp120 differential glycosylation on recognition by broadly neutralizing monoclonal antibodies or by polyclonal antibodies of individuals infected with other HIV-1 subtypes is unknown.
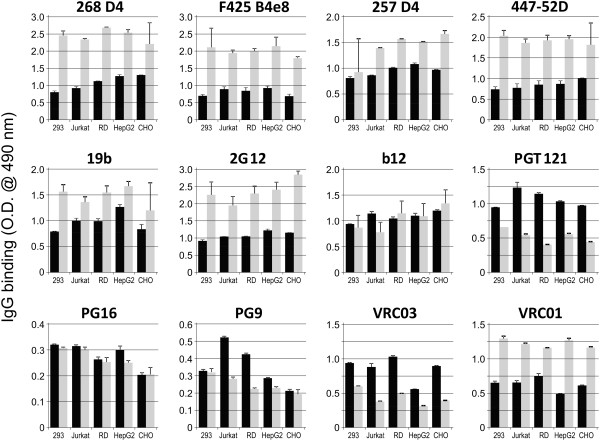
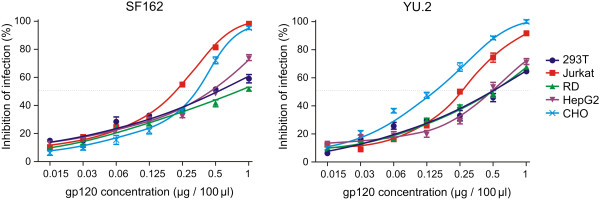
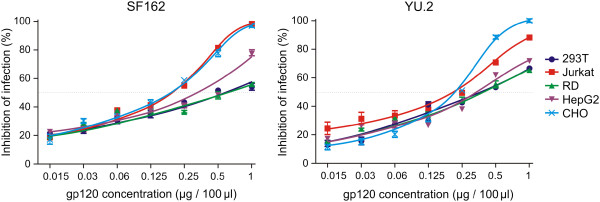
Methods: Recombinant multimerizing gp120 antigens were expressed in different cells, HEK 293T, T-cell, rhabdomyosarcoma, hepatocellular carcinoma, and Chinese hamster ovary cell lines. Binding of broadly neutralizing monoclonal antibodies and polyclonal antibodies from sera of subtype A/C HIV-1-infected subjects with individual gp120 glycoforms was assessed by ELISA. In addition, immunodetection was performed using Western and dot blot assays. Recombinant gp120 glycoforms were tested for inhibition of infection of reporter cells by SF162 and YU.2 Env-pseudotyped R5 viruses.
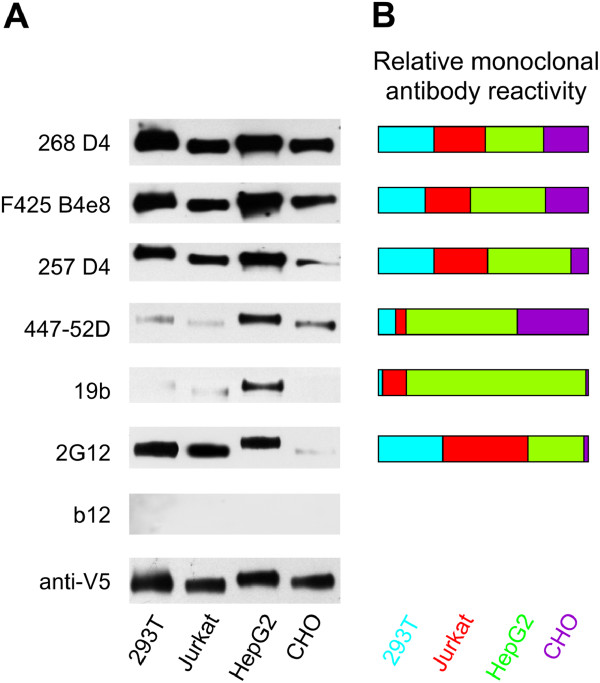
Results: We demonstrated, using ELISA, that gp120 glycans sterically adjacent to the V3 loop only moderately contribute to differential recognition of a short apex motif GPGRA and GPGR by monoclonal antibodies F425 B4e8 and 447-52D, respectively. The binding of antibodies recognizing longer peptide motifs overlapping with GPGR epitope (268 D4, 257 D4, 19b) was significantly altered. Recognition of gp120 glycoforms by monoclonal antibodies specific for other than V3-loop epitopes was significantly affected by cell types used for gp120 expression. These epitopes included CD4-binding site (VRC03, VRC01, b12), discontinuous epitope involving V1/V2 loop with the associated glycans (PG9, PG16), and an epitope including V3-base-, N332 oligomannose-, and surrounding glycans-containing epitope (PGT 121). Moreover, the different gp120 glycoforms variably inhibited HIV-1 infection of reporter cells.
Conclusion: Our data support the hypothesis that the glycosylation machinery of different cells shapes gp120 glycosylation and, consequently, impacts envelope recognition by specific antibodies as well as the interaction of HIV-1 gp120 with cellular receptors. These findings underscore the importance of selection of appropriately glycosylated HIV-1 envelope as a vaccine antigen.
Keywords: Deglycosylation resistance; Glycan-specific antibody; Neutralization inhibition; gp120 glycosylation.
Figures

References
-
- Wyatt R, Kwong PD, Desjardins E, Sweet RW, Robinson J, Hendrickson WA, Sodroski JG. The antigenic structure of the HIV gp120 envelope glycoprotein. Nature. 1998;393:705–711. - PubMed
-
- Pantophlet R, Burton DR. gp120: target for neutralizing HIV-1 antibodies. Annu Rev Immunol. 2006;24:739–769. - PubMed
-
- Wei X, Decker JM, Wang S, Hui H, Kappes JC, Wu X, Salazar-Gonzalez JF, Salazar MG, Kilby JM, Saag MS, Komarova NL, Nowak MA, Hahn BH, Kwong PD, Shaw GM. Antibody neutralization and escape by HIV-1. Nature. 2003;422:307–312. - PubMed
-
- Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, McLaughlin S, Pascuccio M, Gopaul R, McNally J, Cruz CC, Censoplano N, Chung E, Reitano KN, Kottilil S, Goode DJ, Fauci AS. HIV-1 envelope protein binds to and signals through integrin alpha4beta7, the gut mucosal homing receptor for peripheral T cells. Nat Immunol. 2008;9:301–309. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
