

Hepatitis C virus subverts liver-specific miR-122 to protect the viral genome from exoribonuclease Xrn2
- PMID: 25121753
- PMCID: PMC4227615
- DOI: 10.1016/j.chom.2014.07.006
Hepatitis C virus subverts liver-specific miR-122 to protect the viral genome from exoribonuclease Xrn2
Abstract
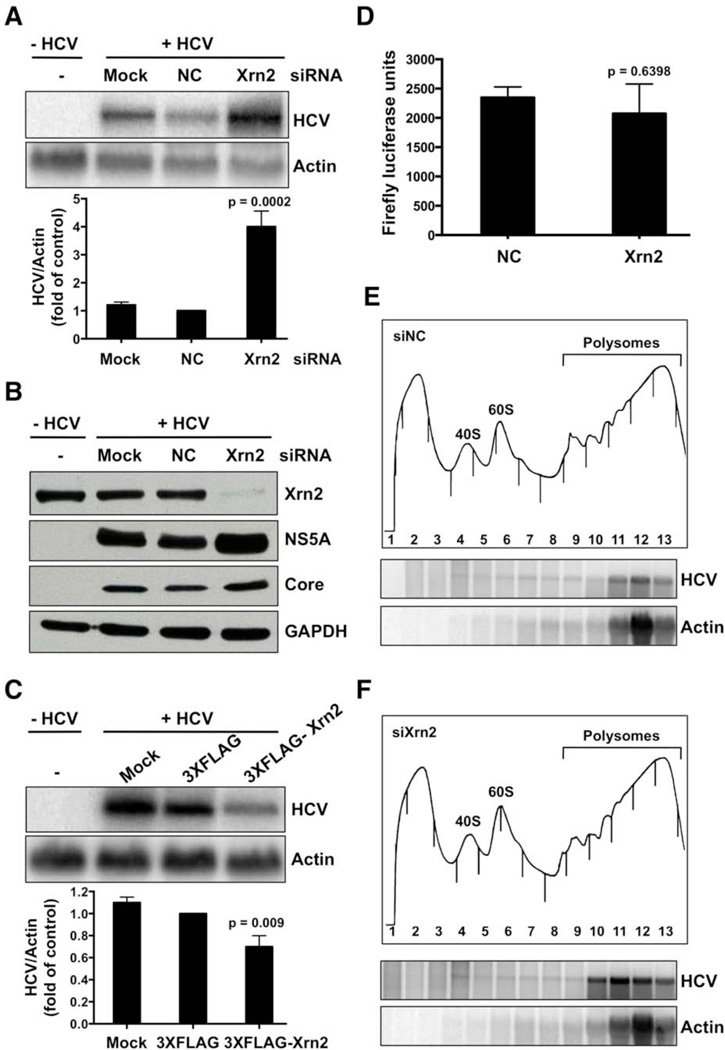
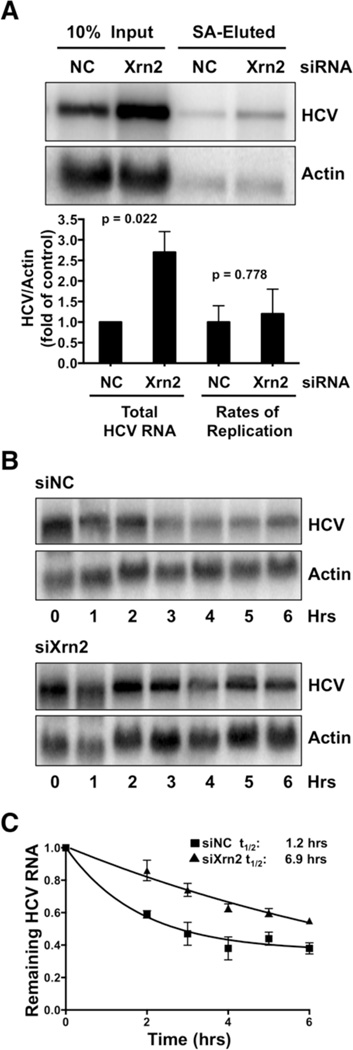
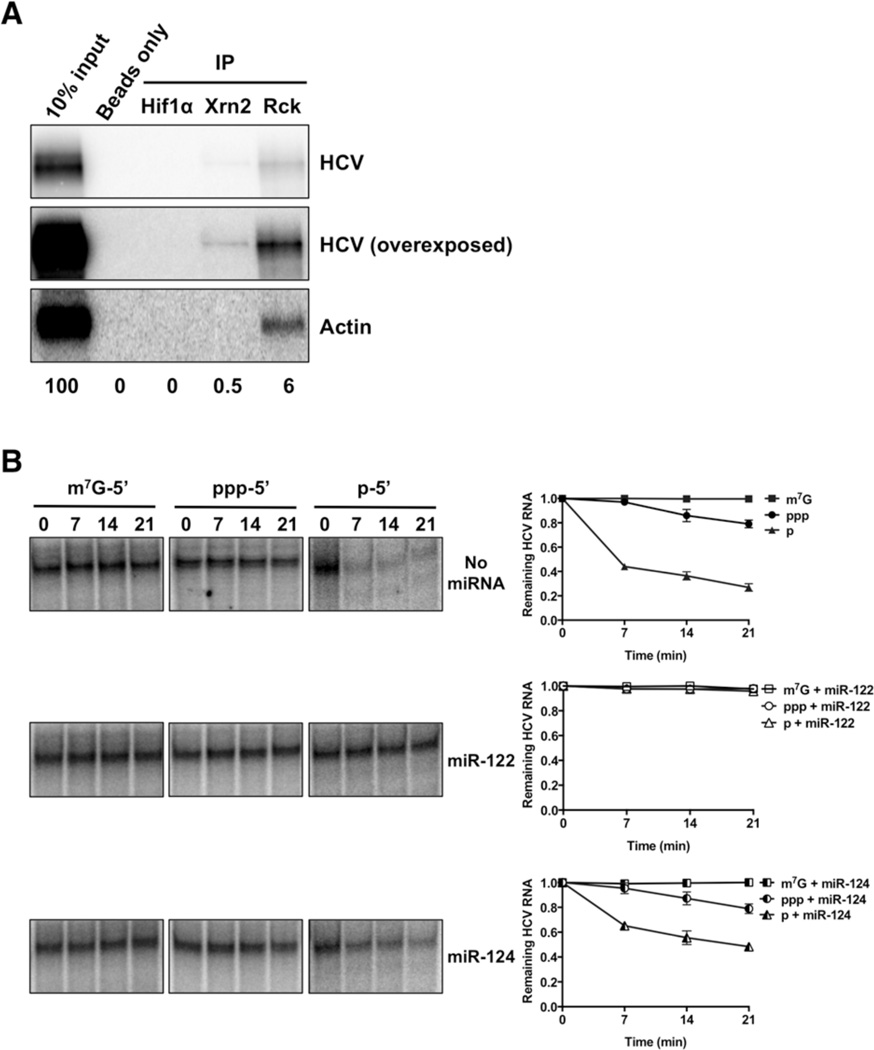
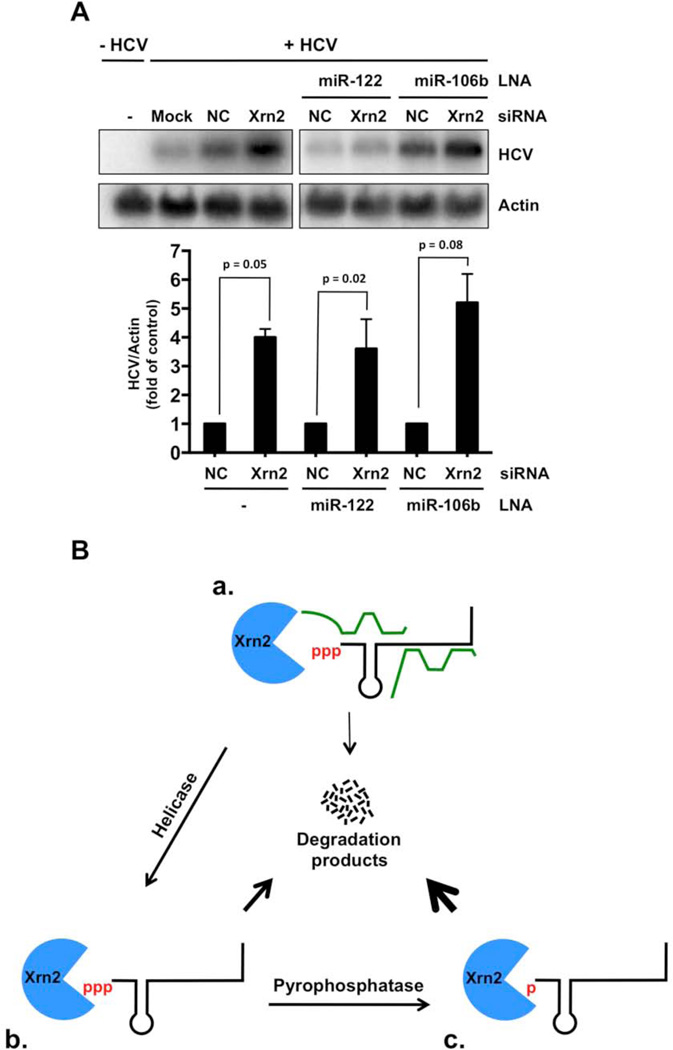
The abundant, liver-specific microRNA miR-122 forms extensive base-pairing interactions with the 5' noncoding region of the hepatitis C virus (HCV) RNA genome, protecting the viral RNA from degradation. We discovered that the 5'-3' exoribonuclease Xrn2, which plays a crucial role in the transcription termination of RNA polymerase II, modulates HCV RNA abundance in the cytoplasm, but is counteracted by miR-122-mediated protection. Specifically, Xrn2 depletion results in increased accumulation of viral RNA, while Xrn2 overexpression diminishes viral RNA abundance. Depletion of Xrn2 did not alter translation or replication rates of HCV RNA, but affected viral RNA stability. Importantly, during sequestration of miR-122, Xrn2 depletion restored HCV RNA abundance, arguing that Xrn2 depletion eliminates the miR-122 requirement for viral RNA stability. Thus, Xrn2 has a cytoplasmic, antiviral function against HCV that is counteracted by HCV's subversion of miR-122 to form a protective oligomeric complex at the 5' end of the viral genome.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Bartenschlager R, Penin F, Lohmann V, Andre P. Assembly of infectious hepatitis C virus particles. Trends in microbiology. 2011;19:95–103. - PubMed
-
- Benoit B, Mitou G, Chartier A, Temme C, Zaessinger S, Wahle E, Busseau I, Simonelig M. An essential cytoplasmic function for the nuclear poly(A) binding protein, PABP2, in poly(A) tail length control and early development in Drosophila. Developmental cell. 2005;9:511–522. - PubMed
-
- Chatterjee S, Grosshans H. Active turnover modulates mature microRNA activity in Caenorhabditis elegans. Nature. 2009;461:546–549. - PubMed
-
- Cleary MD, Meiering CD, Jan E, Guymon R, Boothroyd JC. Biosynthetic labeling of RNA with uracil phosphoribosyltransferase allows cell-specific microarray analysis of mRNA synthesis and decay. Nat Biotechnol. 2005;23:232–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
