

Identification of a sphingolipid α-glucuronosyltransferase that is essential for pollen function in Arabidopsis
- PMID: 25122154
- PMCID: PMC4371831
- DOI: 10.1105/tpc.114.129171
Identification of a sphingolipid α-glucuronosyltransferase that is essential for pollen function in Arabidopsis
Abstract
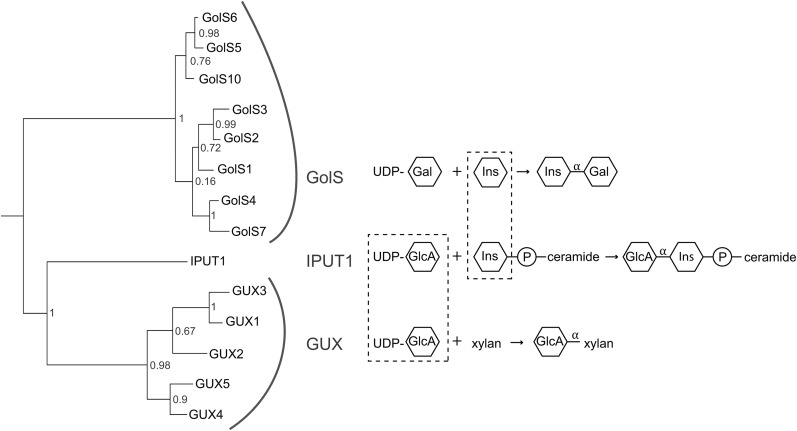
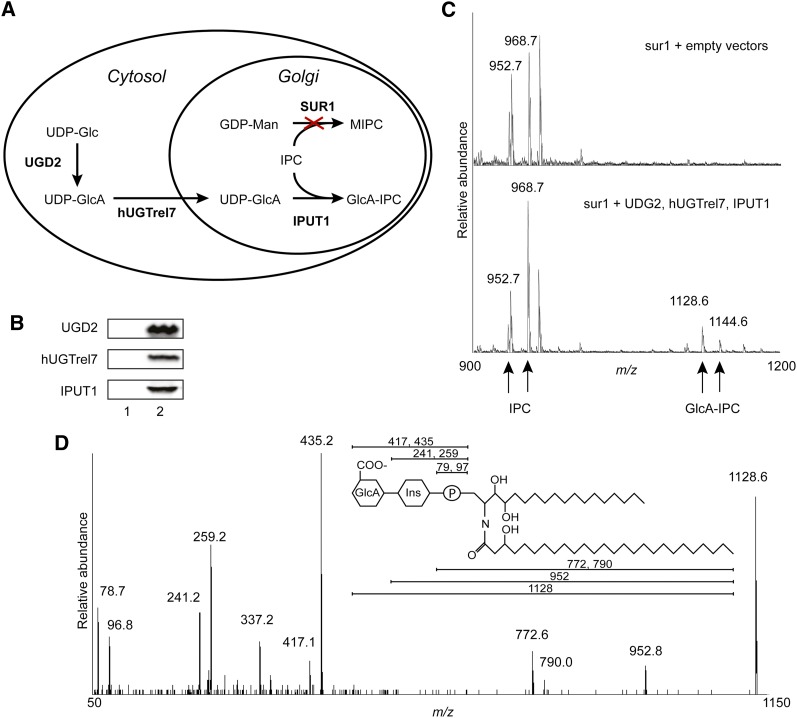
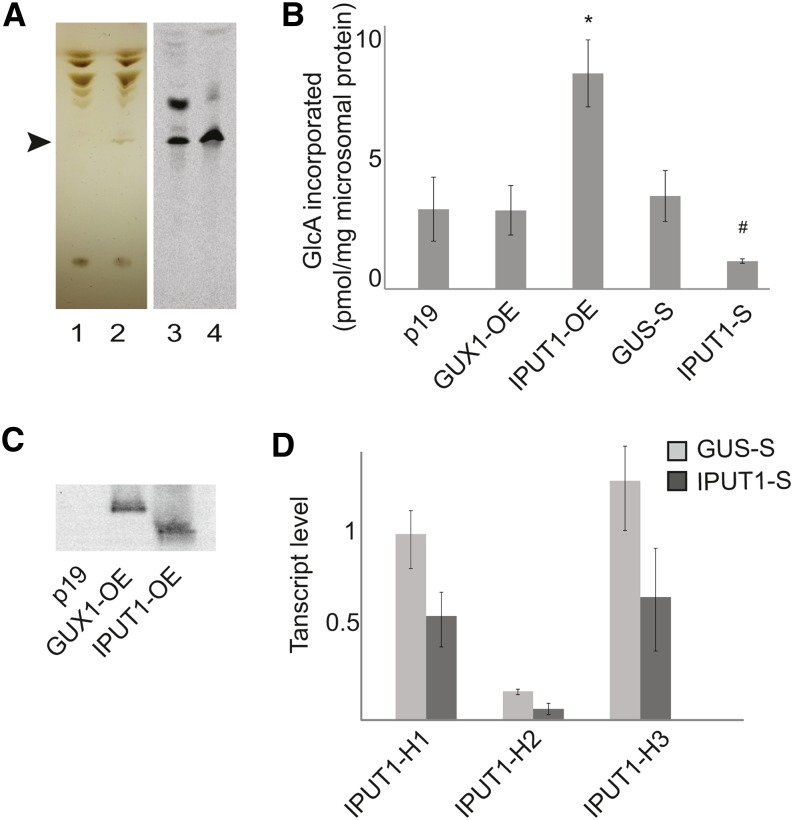
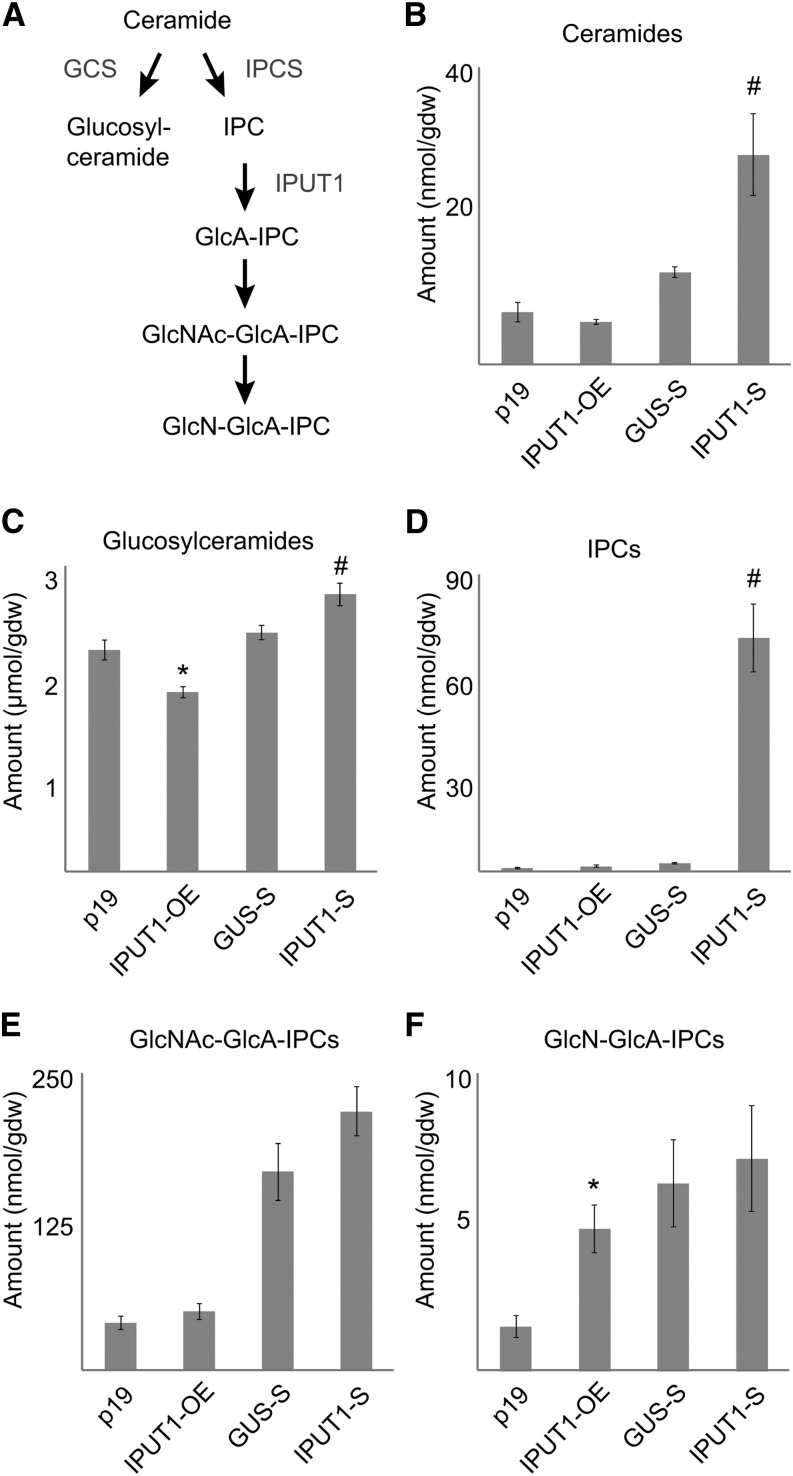
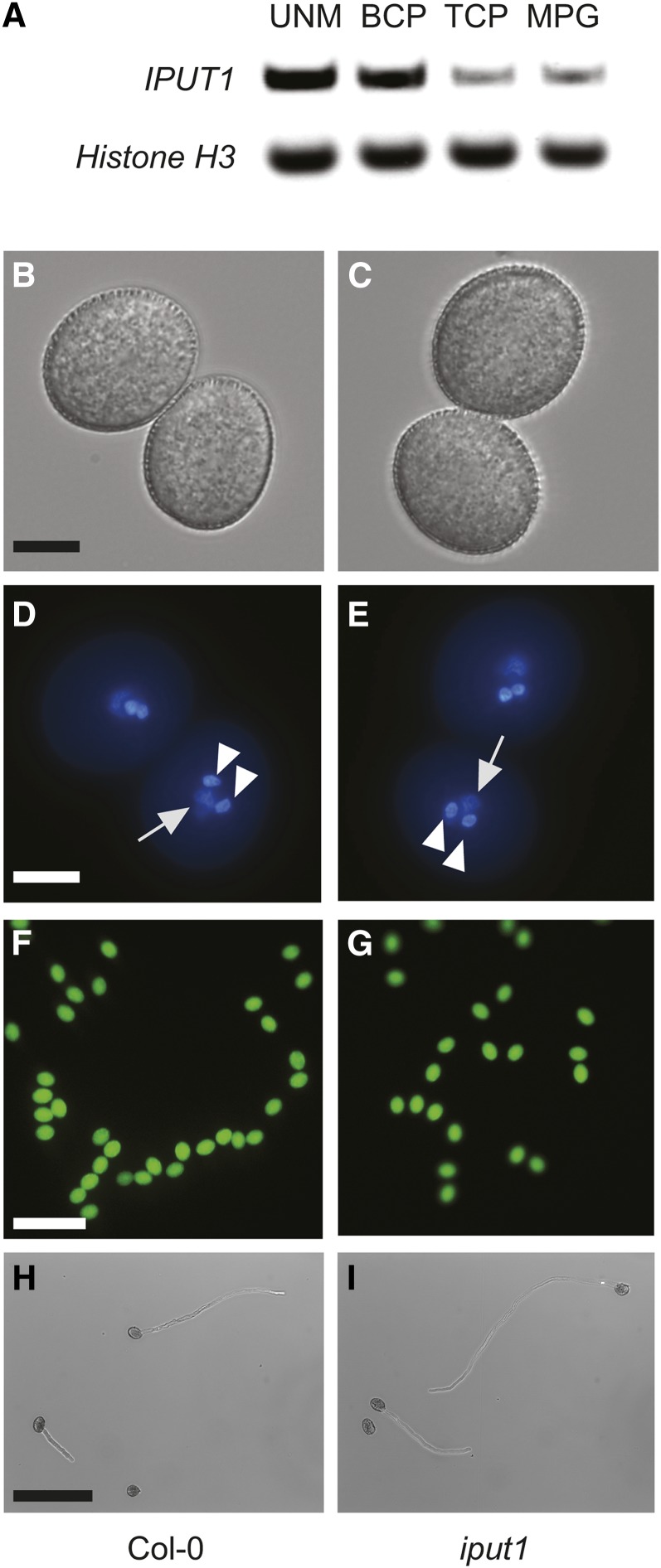
Glycosyl inositol phosphorylceramide (GIPC) sphingolipids are a major class of lipids in fungi, protozoans, and plants. GIPCs are abundant in the plasma membrane in plants, comprising around a quarter of the total lipids in these membranes. Plant GIPCs contain unique glycan decorations that include a conserved glucuronic acid (GlcA) residue and various additional sugars; however, no proteins responsible for glycosylating GIPCs have been identified to date. Here, we show that the Arabidopsis thaliana protein INOSITOL PHOSPHORYLCERAMIDE GLUCURONOSYLTRANSFERASE1 (IPUT1) transfers GlcA from UDP-GlcA to GIPCs. To demonstrate IPUT1 activity, we introduced the IPUT1 gene together with genes for a UDP-glucose dehydrogenase from Arabidopsis and a human UDP-GlcA transporter into a yeast mutant deficient in the endogenous inositol phosphorylceramide (IPC) mannosyltransferase. In this engineered yeast strain, IPUT1 transferred GlcA to IPC. Overexpression or silencing of IPUT1 in Nicotiana benthamiana resulted in an increase or a decrease, respectively, in IPC glucuronosyltransferase activity in vitro. Plants in which IPUT1 was silenced accumulated IPC, the immediate precursor, as well as ceramides and glucosylceramides. Plants overexpressing IPUT1 showed an increased content of GIPCs. Mutations in IPUT1 are not transmitted through pollen, indicating that these sphingolipids are essential in plants.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Beeler T.J., Fu D., Rivera J., Monaghan E., Gable K., Dunn T.M. (1997). SUR1 (CSG1/BCL21), a gene necessary for growth of Saccharomyces cerevisiae in the presence of high Ca2+ concentrations at 37 ° C, is required for mannosylation of inositolphosphorylceramide. Mol. Gen. Genet. 255: 570–579. - PubMed
-
- Boavida L.C., McCormick S. (2007). Technical advance: temperature as a determinant factor for increased and reproducible in vitro pollen germination in Arabidopsis thaliana. Plant J. 52: 570–582. - PubMed
-
- Bromley P.E., Li Y.O., Murphy S.M., Sumner C.M., Lynch D.V. (2003). Complex sphingolipid synthesis in plants: characterization of inositolphosphorylceramide synthase activity in bean microsomes. Arch. Biochem. Biophys. 417: 219–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
